

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfПарная регистрация возможна для идентифицированных нейронов высших животных, включая млекопитающих (рис. 7.14).
5 ìêì
Ðèñ. 7.14. Компьютерная трехмерная объемная реконструкция четырех центральных синапсов с сохранением пропорций в размере (Rollenhagen and Lübke. 2006).
a.Чашечка Хелда — главный нейрон (MNTB), гигантский синапс в медиальном ядре трапецевидного тела в слуховой системе млекопитающих (мышь, крыса, кошка).
b.Синапс между мшистыми волокнами и пирамидными нейронами поля
CA3 гиппокампа.
c-ñ1. Окончания мшистых волокон на дендритах гранулярных нейронов.
d.Аксо-дендритный синапс в новой коре: проходящий аксон — шипик дендрита пирамидной клетки пятого слоя неокортекса.
7.9.1. Идентифицированные синапсы моллюсков
Моллюски являются одним из излюбленных модельных объектов нейробиологов из-за возможности индивидуальной идентификации их нейронов и синапсов (рис. 7.15). Другим преимуществом нейронов моллюсков является их небольшое количество (1–10 тысяч, что на 3–6 порядков меньше, чем у позвоночных), большие размеры клеточных тел, расположение тел нейронов на поверхности ганглиев, их хорошая «выживаемость» (несколько суток на препарате в физиологическом растворе) и возможность внутриклеточной регистрации активности
220
нейронов на препарате, у которого сохранны некоторые функции живого организма («полуинтактные препараты») — все это создало уникальную возможность объединить эксперименты на нейронном уровне с данными поведенческих экспериментов.
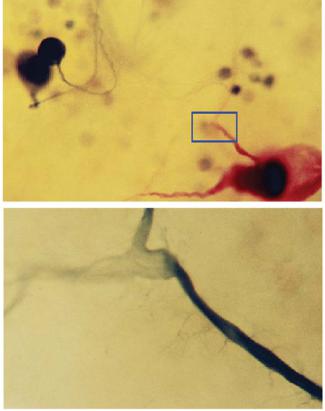
Ðèñ. 7.15. «Простые нервные системы» — нервные системы беспозвоноч- ных животных. Идентифицируемые нейроны и идентифицированные синапсы моллюска, виноградной улитки, Helix pomatia/Helix lucorum). Окрашенные разными методами нейроны левого и правого париетального (на схеме: LPag, RPag) и висцерального (Vg) ганглиев улитки. Красным (внутриклеточно, хлоридом кадмия) окрашен гигантский командный нейрон пассивно-оборонительного рефлекса улитки (LPa2), черным — ретроградное окрашивание нейронов кобальтом через аортальный нерв (Aortal. n.). Внизу — десятикратное увеличение области (выделено синим). Видны ответвления одного из двух основных отростков биполярного командного нейрона. Шкала по горизонтали — 100 мкм.
221
Преимущества морского моллюска, морского зайца, аплизии (Aplysia), были использованы американским нейробиологом Эриком Кэнделом для изучения памяти и научения (рис. 7.16). То, что удалось экспериментально исследовать механизмы научения и памяти на клеточном и молекулярном уровнях, было высоко оценено, и в 2000 году Эрику Кэнделу вместе с Полем Грингардом и Эрвидом Карлсоном была присуждена Нобелевская премия. Рисунки из Нобелевской лекции Э. Кэндела представлены в разделе про синаптическую пластичность.
Ðèñ. 7.16. У моллюска, морского зайца — аплизии (Aplysia) простая нервная система и есть оборонительный рефлекс сокращения жабер (gill). Это было использовано Эриком Кэнделом для изучения механизмов научения и памяти (Рис. из Нобелевской лекции Э. Кэндела).
7.9.2. Идентифицируемые синапсы млекопитающих
Большие размеры синапсов являются значимым преимуществом беспозвоночных животных из-за возможности одновременной внутриклеточной регистрации электрической активности пре- и постсинаптического нейронов (парной регистрации). Это преимущество было использова-
222
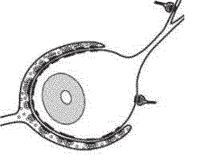
Ðèñ. 7.17. Синапсы Хелда.
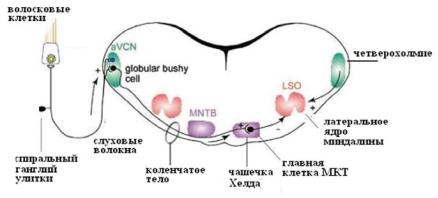
но, что привело к важным открытиям в понимании механизмов функционирования синапсов (Ллинас, 1972; Кэндел, 1980; Kandel, 2000). На позвоночных такие исследования долгое время не были доступны из-за небольших размеров пресинаптических окончаний. В конце 20-го века наше понимание механизмов функционирования синапсов позвоночных животных основывалось на данных, полученных на нескольких модельных объектах, таких, как нервно-мышесное соединение (Кац, Katz B., 1972), хромаффинные клетки цилиарного ганглия цыпленка (Роуз, 1995), нервные окончания в нейрогипофизе, синаптосомальные препараты (препараты изолированных клеток в культуре ткани). В ЦНС млекопитающих большая часть синаптических соединений является быстрыми глутаматергическими синапсами. Но долгое время такой препарат с глутаматергическими нейронами, от пресинаптических терминалей которых можно отводить внутриклеточную регистрацию с фиксацией потенциала (patch clamp), был только предметом мечтаний. Одним из кандидатов на роль такого препарата были мшистые волокна в гиппокампе, другим — гигантские синапсы, обнаруженные морфологами в слуховой системе ствола мозга: «бутоновидные окончания» Хелда (the enddulbs of Held) и чашечки Хелда (the calyxes of Held). Оба синапса являются глутаматэргическими и расположены в проводящих путях слуховой системы, обеспечивающих локализацию источника звука (рис. 7.17). Своим названием они обязаны немецкому анатому Гансу Хелду, исследовавшему в конце 19-го века центральные пути слуховой системы кошки (Held, 1893). Используя метод окрашивания по Гольджи, Хелд обнаружил два типа гигантских синапсов, получивших впоследствии его имя: окончание Хелда и чашечка Хелда.
223
Эти синапсы с большим энтузиазмом изучал Сантьяго Рамон-и-Ка- хал (Кахал, 1985). Он получил первое изображение синапсов Хелда с большим разрешением с помощью светового микроскопа (рис. 7.18).
Ðèñ. 7.18. Синапс Хелда. Рисунок одной чашечки Хелда на одном главном нейроне медиального ядра коленчатого тела (MNTB). Клетки MNTB получают добавочные тормозные и возбуждающие входы через небольшие бутоновидные синапсы, но в большинстве случаев одна главная MNTB клетка получает вход только через один огромный чашечковидный синапс Хелда.
Выводы
•Основной тип связи между нервными клетками в мозге — это синаптическая связь. Основной тип синаптической связи — химические синапсы. Химическая передача сигналов осуществляется путем экзоцитоза пресинаптическими терминалями химических посредников
èвоздействия нейромедиаторов на рецепторы мембраны постсинапти- ческой клетки.
•В нервной системе существуют и несинаптические способы хими- ческой передачи сигналов, например, все большее внимание привлекает объемная передача и спиловер, «утечка» медиатора. Электрические синапсы существуют в мозге в виде исключения.
•Многообразие синапсов в нервной системе сравнимо с многообразием нейронов. Синапсы различаются по размеру (в сотни раз), по функциональным и динамическим параметрам. Химические синапсы разли- чаются по эргичности — медиатору, передающему сигнал. Структурные
èхимические различия синапсов определяют их функциональные и динамические различия, такие, как сила связи, скорость проведения сигнала и синаптическая задержка, знак и эффективность воздействия. Динамические параметры синапсов определяют их пластичность.
224
•При всем многообразии конкретных видов и типов синапсов общие принципы их функционирования удивительно сходны на всех уровнях эволюционного развития, от беспозвоночных до высших позвоноч- ных животных. Сходство механизмов синаптической передачи сигналов и ее пластичности наблюдается и на клеточном, и на молекулярном уровнях.
•Динамические параметры передачи сигнала могут быть смоделированы на основе данных, полученных на идентифицируемых синапсах, а результаты моделирования проверены на препаратах нервной системы и в поведенческих психофизиологических экспериментах. Компьютерное моделирование синапсов стало необходимым звеном клеточных и молекулярно-генетических исследований механизмов работы мозга и психофизиологических исследований.
Цитированная литература
Рекомендуемая
1.Николлс Дж. Г., Мартин А. Р., Валлас Б. Дж., Фукс П. А. От нейрона к мозгу М.: Изд. УРСС, 2003, 672 с.
2.Соколов Е. Н. Восприятие и условный рефлекс: новый взгляд. М.: УМК «Психология», 2003, 287 с.
Дополнительная
1.Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекуляpная биология клетки. М.: Миp. 1994, т. 2, гл. 12, с. 338-383; ò. 3, ãë. 16: ñ. 67, 78, 81, 94-100, ãë. 19: ñ. 287-375.
2.Анохин К. В. Молекулярные сценарии консолидации долговременной памяти // Журн. высш. нервн. деят., 1997. т. 47, ¹2, с. 261-286 (Хрестоматия по физиологии ВНД, УМК «Психология», Москва 2000, с. 217-230).
3.Анохин П. К. (ред.) Системный анализ интегративной деятельности нейрона. М.: Наука, 1974. 158 с.
4.Ашмарин И. П. Загадки и откровения биохимии памяти / Под ред. акад.
Å.М. Кребса. Л.: Изд-во Ленинградского Университета, 1975, 160 с.
5.Ашмарин И. П., Кругликов Р. И. Пептиды, обучение, память (принцип полифункциональности). Нейрохимия, 1983, т. 2. ¹3. С. 327.
6.Блум Ф., Лейзерсон А., Ховстедтер Л. Мозг, разум и поведение. М.: Мир, 1988. 64 с.
7.Балабан П. М., Захаров И. С. Обучение и развитие: общая основа двух явлений. М.: Наука, 1992. 152 с.
7.Виноградова О. С. (2000) Нейронаука конца второго тысячелетия: смена парадигм // Журн. высш. нервн. деят. 2000. Т. 50. ¹50. С. 743-774.
225
8.Гомазков О. А. Нейротрофическая регуляция и стволовые клетки мозга. М.: Èçä-âî ÈÊÀÐ, 2006, 332 ñ.
9.Гомазков О. А. Мозг и нейропептиды. 1997; Нейропептиды и ростовые факторы мозга. 2002; Нейрохимия возрастных ишемических заболеваний мозга. 2003.
10.Греченко Т. Н. Нейро-физиологические механизмы памяти. М.: Наука, 1979. 165 с.
11.Кибяков А. И., Сахаров Д. А. Рассказы о медиаторах. М.: Знание, 1978. 64 с.
12.Куффлер С., Николс Дж. От нейрона к мозгу. М.: Мир. 1979.
13.Кэндел Э. Клеточные основы поведения. М.: Мир, 1980. 598 с. Kandel E. R. (2000) The Nobel Prize in Physsiology or Medicine 2000. (Kandel_Nobel lecture_2000.ppt).
14.Окс С. Основы нейрофизиологии. М.: Мир, 1968. Гл. 1-8.
15.Первис Р. Микроэлектродные метолы внутриклеточной регистрации и ионофореза. М.: Мир, 1983. Регистрация от одиночных ионных каналов. М.: Мир, 1987.
16.Прибрам К. Языки мозга. М.: Мир, 1975. С. 17-64.
17.Роуз С. Устройство памяти: от молекул к сознанию М.: Мир, 1995. 380 с.
18.Сахаров Д. А. Генеалогия нейронов. М.: Наука, 1974. 183 с.; Сахаров Д. А. Медиаторная специфичность нейронов: происхождение и эволюция. М., 1973. 46 с.
19.Сахаров Д. А. (1990) Множественность нейротрансмиттеров: функциональное значение. Журн. эвол. биохим. и физиол., 1990, т. 26, ¹5, с. 733-749.
20.Свердлов Е. Д. Очерки современной молекулярной генетики. // Молек. генет. микробиол. и вирусолог. 1995, ¹¹ 2, 3, 4, 5, 1996 ¹5, 1997 ¹4.
21.Сентаготаи Я., Арбиб М. Концептуальные модели нервной ткани. М.: Мир, 1976. 198 с.
22.Соколов Е. Н., Шмелев Л. А. Нейробионика. М.: Наука, 1983.
23.Соколов Е. Н. Нейронные механизмы памяти и научения. М.: Наука, 1981. 181 с.
24.Сторожук В. М. Нейронные механизмы обучения. Киев: Наук. Думка, 1986. 264 с.
25.Шаповалов А. И., Ширяев Б. И. Передача сигналов в межнейронных синапсах. Л.: Наука, Лен. отд., 1987. 173 с.
26.Шеперд Г. Нейробиология: В 2-õ ò. Ì.: Ìèð, 1987. 454 ñ.
27.Шмидт Р. и Тевс Г. (Ред.) Физиология человека. М.: Мир, 1996. В 4-õ ò.
28.Эделман Дж., Маунткасл В. Разумный мозг. М.: Мир, 1981. 135 с.
29.Basic Neurochemistry. 6th Edition. Eds.: G. J.Siegel, B. W. Agranoff, R. W. Alberts, S. K. Fisher, M. D. Uhler. Part two: Intercellular signaling.
30.Cell and Tissue Research/ September. Biomedical and Life Sciences and Medicine. Springler Berlin / Heidelberg 23, 2006, v.326, pp. 201-669.
31.Klaus Unsicker (from the Editor:) The synapse — recent advances. P. 201;
32.Frotscher M., Gundelfinger E., Jonas P., Neher E., Seeburg P. (Editorial) The most important advances in synapse research from my point of view — and what remains to be done. P. 203-204.
226
33.Jan L. Gibbins and Judy L. Morris Structure of peripheral synapses: autonomic ganglia. Review, pp. 205-220.
34.Astrid Rollenhagen and Joachim H.R. Lübke The morphology of excitatory central synapses: from structure to function. (Review), pp. 221-237.
35.Scheggenburger, Ralf and Forsythe, Jan D. The Calyx of Held. (Review), pp. 311-337.
36.Анохин П. К. Узловые вопросы теории функциональной нервной системы. М.: Наука, 1980, 196 с. (Петр Кузьмич Анохин (1898-1974). М.: Наука, 1987. 128 с.
37.Виноградова О. С. Гиппокамп и память. М.: Наука, 1975. 333 с.
38.Кахаль С. Р. Автобиография (воспоминания о моей жизни) / Под ред. А.В. Смольянникова и Д.С. Саркисова. Пер. с англ. М.: Медицина, 1985. 272 с.
39.Êàö Á. (Katz B.) The release of the neural transmitter substances. Liverpool University Press, Liverpool, 1969.
40.Павлов И. П. Избранные труды / Ред. Э. А. Асратян. М.: Изд-во Академии педагогических наук РСФСР, 1951. 616 с.
41.Павлов И. П. (1926, 1927, 1935, 1952) Лекции о работе больших полушарий головного мозга. Москва: Академия медицинских наук СССР, 1952. 287 с.
42.Сеченов И. М. Рефлексы головного мозга: Попытка свести способ происхождения психических явлений на физиологические основы. С биографией И. М. Сеченова (М. Н. Шатерников, с. 5-24). М.: Комкнига (Из наследия мировой психологии), 2007, 127 с.
43.Соколов Е. Н. Восприятие и условный рефлекс. М.: Издательство МГУ им. М. В. Ломоносова, 1958.
44.Соколов Е. Н. Механизмы памяти (опыт экспериментального исследования). Изд-во Московского Университета, 1969. 176 с.
45.Сомьен Дж. Кодирование сенсорной информации (George Somjen, 1972. Sensory Coding in the mammalian nervous system). М.: Мир, 1975. 415 с.
46.Тасаки И. Проведение нервного импульса. М.: Иностранная литература, 1957. 187 с.
47.Тасаки И. Нервное возбуждение. М.: Мир, 1971. 222 с.
48.Теория химической передачи нервного импульса (этапы развития). Сост. Михельсон М. Я. Л.: Наука, Ленинградское отд., 1981. 144 с.
49.Ходжкин А. Нервный импульс. М.: Мир, 1965, 125 с. (Hodgkin A.L. The Conduction of the Nervous Impulse/Liverpool University Press, 1964).
50.Экклс Дж. Физиология нервных клеток. М.: Иностранная литература, 1959. 298 с.
51.Экклс Дж. Физиология синапсов. М.: Мир, 1966. 395 с. (Перевод с англ. О.С. Виноградовой. Под ред. и с предисловием П. К. Анохина) / John Carew Eccles (1964) The physiology of synapses. Spriner-Verlad, Berlin, Gettingen, Heldelberg.
52.Hebb D. O. (McGill Univ.) The Organization of Behavior/ A Neuropsychological Theory. USA, N.Y., John Wiley & Sons, Ink.; London, Charman & Hall, Limited, 1949, P. 335.
53.Sir John Eccles — Nobel Lecture, December 11, 1963: The Ionic Mechanism of Postsynaptic Inhibition.
227
Глава 8. СИНАПТИЧЕСКАЯ ПЛАСТИЧНОСТЬ
В главе рассмотрены механизмы пресинаптической и постсинаптической пластичности. В основе пресинаптической пластич- ности лежат внутриклеточные каскады, инициируемые серотонином в пресинаптическом нейроне. Пресинаптическая пластич- ность определяется усилением (или уменьшением) выделения медиатора из пресинаптического нейрона. Постсинаптическая пластичность связана с изменением состояния и числа рецепторов постсинаптической мембраны. Кратковременная пластич- ность обусловлена процессами, происходящими в мембранах синаптических контактов. При долговременных модификациях синаптической передачи дополнительно инициируются процессы экспрессии генов с последующим синтезом различных белков-ре- гуляторов, факторов роста активных зон и синапсов в пресинаптических мембранах и синтезом мембранных рецепторов в постсинаптических мембранах. Синаптическая пластичность рассматривается как клеточная основа памяти и обучения.
8.1. Типы синаптической пластичности
Синаптическая пластичность заключается в изменении эффективности синаптической передачи в результате интенсивной ритмической стимуляции афферентных путей нейрона. В результате такая стимуляция может либо увеличивать (синаптическая сенситизация, фасилитация, облегчение, потенциация), либо уменьшать (синаптическая депрессия) результирующие постсинаптические потенциалы (ПСП) в постсинаптическом нейроне.
По временному критерию изменение эффективности может быть кратковременным (сотни миллисекунд, секунды, минуты) и долговременным (часы и дни). К кратковременным эффектам относят фасилитацию, усиление и кратковременную депрессию. Относительно мощный высокочастотный залп стимулов (тетанизация) вызывает более продолжительный эффект — посттетаническую потенциацию, которая заключается в увеличении амплитуды ПСП в течение десятков минут. Тетанизация может вызывать и более длительное усиление ПСП в нейронах, называемое долговременной потенциацией (ДВП), или ослабление ПСП, называемое долговременной депрессией (ДВД), продолжающееся часы и даже дни.
228
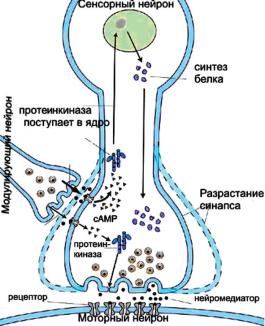
По механизму такие изменения могут быть локализованы в пресинаптическом и постсинаптическом нейронах. Многообразие пресинаптических механизмов заключается, в конечном счете, в увеличении выброса медиатора из пресинаптического волокна. Постсинаптические механизмы приводят к увеличению чувствительности постсинаптической мембраны к медиатору— сенситизации рецепторов (усилению эффективности связывания с медиатором или увеличению проводимости в канале) — и увеличению числа рецепторов.
Современные модели синаптической пластичности основаны на результатах, полученных при изучении нейронной организации примитивного поведения моллюсков и феномена длительной потенциации на переживающих срезах гиппокампа и коры млекопитающих.
8.2. Пресинаптическая пластичность
Механизмы пресинаптической пластичности изучаются на относительно простых нейронных моделях моллюсков — аплизии (Aplysia) и виноградной улитки (Helix lucorum). Эти животные имеют относительно простую нервную систему из нескольких ганглиев, состоящих из больших (до 1 мм), легко идентифицируемых нейронов. Такая простая нервная система является удобным объектом для анатомических, биофизических и биохимических исследований. Примитивное поведение этих животных обеспечивается активностью нескольких нейронов, образующих простую нервную сеть. Некоторые примитивные поведен- ческие акты могут модифицироваться в результате обучения. При этом участки в нервной сети, которые обеспечивают эти модификации, могут быть идентифицированы. Поэтому нейронные механизмы, лежащие в основе модификаций поведения моллюсков, поддаются экспериментальному анализу и моделированию.
8.2.1.Кратковременная фасилитация сенсомоторных связей
âнервной системе моллюсков
Не вдаваясь в подробности поведенческих экспериментов, рассмотрим некоторые феномены, которые отражают изменения в постсинаптических ответах мотонейронов, обеспечивающих простые поведенческие реакции моллюсков. Поведенческие ответы аплизии обеспечиваются простой нейронной сетью, состоящей из нескольких нейронов (рис. 8.1). Механическое или электрическое раздражение кожи сифона вызывает активацию сенсорного нейрона, аксонная коллатераль которого оканчивается на мембране моторного нейрона (рис. 8.1, снизу).
229