

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
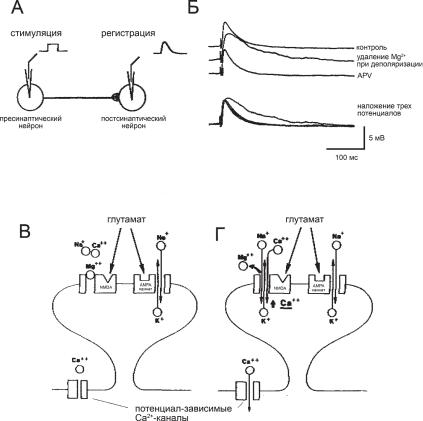
.pdf-30 мВ, при котором Ма2+-блок снимается и канал пропускает смешанный Na+/K+/Ca2+-ток, вызывающий более продолжительный ВПСП (рис. 8.8, Б, удаление Mg2+, Г). NMDA–рецептор остается активированным в течение длительного времени, обеспечивая продолжительную деполяризацию. Сами NMDA-рецепторы обладают относительно высокой Са2+-проводимостью и при активации повышают концентрацию Са2+ в нейроне. Кроме того, значительная деполяризация, обеспечиваемая NMDA–рецепторами, активирует потенциал-зависимые Са2+-каналы в мембране нейрона, в результате чего в нейроне еще более повышается концентрация Са2+ (рис. 8.8, Г). Специфический блокатор NMDA-ре- цепторов APV устраняет продолжительную активацию (рис. 8.8, Б). Участие NMDA-рецепторов в механизме ДВП подтверждено экспериментами, в которых антагонист NMDA-рецепторов APV блокирует запуск ДВП.
Ñà2+-механизм индукции ДВП подтверждается также исследованиями, в которых показано устранение ДВП при внутриклеточном введении буферов, блокирующих увеличение градиента Са2+. Кроме того, поток Са2+ из внутриклеточных депо также может вызывать эффект ДВП. Внутриклеточный Са2+ в повышенной концентрации может активировать целый ряд внутриклеточных каскадов. Наиболее существенными для ДВП является активация СаМКII и цАМФ-зависимой протеинкиназы. Так, внутриклеточная инъекция блокаторов СаМКII и цАМФзависимой протеинкиназы блокирует индукцию ДВП.
8.3.4. Молекулярные механизмы ДВП
Первоначально считали, что проявление ДВП обусловлено пресинаптическим механизмом, то есть увеличением выделения глутамата из пресинаптических терминалей, как это происходит при кратковременных пластических изменениях — при фасилитации, усилении и постетанической потенциации синаптической передачи. В ряде экспериментов было показано увеличение среднего квантового содержания глутамата при активации терминали после тетанизации, хотя в других исследованиях показано, что при этом происходит не увеличение числа квантов, а увеличение квантового ответа, вызванного увеличением амплитуды ВПСП (постсинаптический механизм). В дальнейшем был сделан ряд открытий, подтверждающих постсинаптический механизм ДВП.
Регуляция количества синаптических рецепторов.
В настоящее время существуют доказательства того, что при индукции ДВП число АМРА-рецепторов в постсинаптических мем-
240
бранах увеличивается. С использованием флюоресцентных красителей было показано встраивание новых меченых рецепторов в мембраны дендритных шипиков. Высказываются предположения о том, что большое число шипиков являются «молчащими» и при тетанизации в их мембрану встраиваются новые АМРАрецепторы, которые приводят к увеличению постсинаптических ответов.
Ðèñ. 8.8. Особенности активации NMDA-рецепторов. (А) Схема регистрации. (Б) ВПСП нейрона при активации глутаматэргического входа в различ- ных условиях. (В) Ионная проводимость при слабой активации возбудительного входа. (Г) Ионная проводимость при сильной активации возбудительного входа. Объяснения в тексте.
241
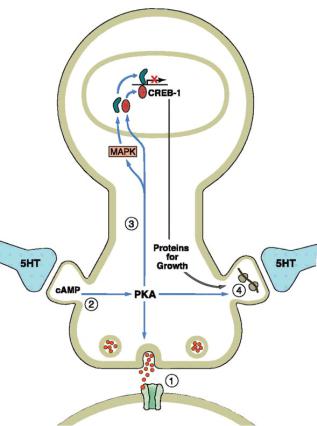
Ðèñ. 8.9. Диалог между генами и синапсами (Кэндел, 2000).
1.Медиатор открывает лиганд-зависимые каналы, что приводит к активации постсинапса в течение нескольких миллисекунд.
2.Активация медиатором трансмембранных рецепторов и включение киназы как вторичного посредника приводит к синаптической активации постсинапса на протяжении нескольких минут.
3.Повторная активация нейротрансмиттером трансмембранных рецепторов приводит к перемещению киназы к ядру клетки и к запуску механизмов генетической транскрипции, что, в конечном счете, приводит к синтезу новых белков, в том числе новых рецепторов медиатора, что обеспечивает долговременное повышение эффективности синаптической передачи.
4.Активация местного синтеза белка, обеспечивает повышение эффективности синаптической передачи в конкретном синапсе, принявшем участие в проведении сигнала.
242
Синтез новых рецепторов.
Методами имунногистохимии показано, что синапсы, образованные коллатералями Шаффера и комиссуральными волокнами на нейронах СА1, содержат NMDA рецепторы, и только часть их содержит АМРА-рецепторы. ДВП приводит к встраиванию в мембраны вновь синтезированных АМРА-рецепторов.
Подтверждением тому являются данные, что ДВП уменьшается при инъекции в постсинаптическую клетку агентов, препятствующих слиянию мембран. Встраивание новых рецепторов в мембрану происходит в результате слияния мембранных транспортных систем (везикул с новыми рецепторами) с клеточной мембраной. Это приводит к увели- чению числа синапсов и, как было показано, к увеличению плотности шипиков (Рис. 8.9).
Ðîëü Ñà2+. Высокая деполяризация после тетанизации в постсинаптических нейронах приводит к увеличению внутриклеточной концентрации Са2+. Ñà2+ связывается с кальмодулином, и комплекс Са2+/ кальмодулин активирует СаМКII, которая автофосфорилируется и сохраняет активность в течение долгого времени, даже после восстановления концентрации Са2+ на начальном уровне. СаМКII оказывает эффекты на синаптическую передачу: 1) фосфорилирует АМРА-рецепторы в мембране, увеличивая их чувствительность и проводимость для ионов, способствуя таким образом усилению ответа рецептора; 2) облегчает мобилизацию (рециклирование) пула резервных АМРА-рецепторов из цитоплазмы в плазматическую мембрану, что приводит к увеличению доступных для активации глутаматом рецепторов.
Относительно кратковременные изменения синаптической пластичности (до 3 час.) происходят за счет активации Са2+- зависимых киназ и протеаз, которые обеспечивают фосфорилирование мембранных канальных белков и белков цитоскелета, ответственных за рециклизацию AMPA-рецепторов из цитоплазматического пула. Длительные эффекты (более 3 час.) происходят за счет активация белкового синтеза. Са2+ активирует Са2+-зависимую аденилатциклазу, затем активированная цАМФ протеинкиназа А фосфорилирует регуляторный элемент (CREB), который, проникая в клеточное ядро, запускает синтез новых глютаматных рецепторов.
243
8.4. Пресинаптическая и постсинаптическая депрессия
Пресинаптическая и постсинаптическая депрессия, так же, как и потенциация, связана с входом Са2+ в клетку. Предполагается, что высокая концентрация Са2+ приводит к ДВП, а умеренная — к ДВД. Такое различие в эффектах Са2+ достигается за счет активации разных Ñà2+-зависимых каскадов как следствие фосфорилирования и дефосфорилирования функционально разнообразных внутриклеточных белков. При долговременной депрессии активируются фосфатазы и кальцинейрин, противодействующие эффекту протеинкиназ. В постсинаптическом нейроне это приводит к уменьшению числа глутаматных АМРА-рецепторов в мембране èç-çà их эндоцитоза (интернализации) — захват из мембраны в цитоплазму.
Выводы
•Синаптическая пластичность является базовым механизмом памяти и обучения. Типы синаптической пластичности различаются по длительности изменений (кратковременная и долговременная пластич- ность), по знаку изменений (фасилитация, сенситизация, потенциация
èдепрессия) и по участию «третьего входа» (гомо- и полисинаптическая и ассоциативная).
•Механизмом кратковременной пресинаптической пластичности является увеличение количества выделяющегося медиатора из сенсорного нейрона в синапсе на командном нейроне.
•Синаптические механизмы активации генома через систему вторичных посредников рассматриваются как основной механизм выбора между кратковременными и долговременными формами пластичности. Долговременная пресинаптическая пластичность заключается в росте активных зон пресинаптической мембраны и развитии новых синапти- ческих контактов.
•В результате высокочастотной стимуляции синапсы в некоторых участках нервной системы увеличивают свою эффективность (посттетаническая потенциация), выражающуюся в увеличении амплитуды синаптических ответов по сравнению с ответами до высокочастотной стимуляции.
•Предполагаемым механизмом индукции посттетанической потенциации является увеличение концентрации внутриклеточного Са2+ в постсинаптическом нейроне, вызванное активацией глутаматных NMDA-
244
рецепторов. Эти рецепторы поддерживают продолжительную деполяризацию, обеспечивая продолжительные Са2+-токи, способствующие усилению синаптического ответа.
•Ðîëü Ñà2+ в поддержании потенциации заключается в том, что в комплексе с кальмодулином он активирует Са2+/кальмодулин-зависи- мую протеинкиназу II. Эта киназа (1) фосфорилирует АМРА-рецепто- ры в мембране, увеличивая их чувствительность и проводимость для ионов, а также (2) облегчает мобилизацию пула резервных АМРАрецепторов из цитоплазмы в плазматическую мембрану.
•Другой предполагаемый механизм заключается в том, что Са2+ запускает механизм внутриклеточной сигнализации, приводящий к фосфорилированию регуляторного элемента CREB, который, проникая
âклеточное ядро, запускает синтез новых глютаматных рецепторов.
•При всем многообразии конкретных видов и типов синапсов общие принципы их функционирования сходны на всех уровнях эволюционного развития, от беспозвоночных до высших позвоночных животных. Сходство механизмов синаптической передачи нервных сигналов и ее пластичности отмечается как на клеточном, так и на молекулярном уровнях.
Цитированная литература
Рекомендуемая
1.Николлс Дж. Г., Мартин А. Р., Валлас Б. Дж., Фукс П. А. От нейрона к мозгу. М.: Изд. УРСС, 2003. 672 с.
2.Соколов Е. Н. Восприятие и условный рефлекс: новый взгляд. М.: УМК «Психология», 2003. 287 с.
Дополнительная
1.Анохин К. В. Молекулярные сценарии консолидации долговременной памяти.Журн. высш. нервн. деят., 1997. Т. 47, ¹2, с. 261-286 (Хрестоматия по физиологии ВНД, УМК «Психология», М., 2000. С. 217-230).
2.Балабан П. М., Захаров И. С. Обучение и развитие: общая основа двух явлений . М.: Наука, 1992. 152 с.
3.Греченко Т.Н. Нейрофизиологические механизмы памяти. М.: Наука, 1979. 165 с.
4.Кэндел Э. Клеточные основы поведения. М.: Мир, 1980. 598 с.
5.Kandel E. R. The Nobel Prize in Physiology or Medicine 2000. (Kandel_Nobel lecture_2000.ppt).
6.Роуз С. Устройство памяти: от молекул к сознанию. М.: Мир, 1995. 380 с.
7.Соколов Е. Н. Нейронные механизмы памяти и научения. М.: Наука, 1981. 181 с.
245
8.Экклс Дж. Физиология синапсов. М.: Мир, 1968.
9.Kandel E. R., Schwartz J. H., Jessell Th. M. Essentials of neural science and behavior Appleton & Lange, Stamford, 1995.
10.The synaptic organization of the brain. Ed. by M. Shepperd, 4th ed., Oxford University Press, NY, Oxford, 1998.
11.Zigmond M. J., Bloom F. E., Landis S. C., Roberts J. LSquire., L. R. Fundamental Neuroscience. Academic Press, San Diego, London, 1999.
246
Глава 9. НЕЙРОГЕНЕТИЧЕСКИЙ МЕХАНИЗМ ФОРМИРОВАНИЯ СЛЕДА ДОЛГОВРЕМЕННОЙ ПАМЯТИ
Нейроны относятся к числу самых пластичных клеток организма. Можно сказать, что процессы развития в мозге никогда не прекращаются. Однако, в отличие от эмбрионального развития, долговременные клеточные перестройки во взрослом мозге находятся под контролем системных когнитивных процессов. В настоящей главе рассматриваются некоторые из известных нейрогенетических механизмов, обеспечивающих долговременное хранение памяти, а также пути их регуляции во время обучения.
9.1.Память у человека способна храниться
âтечение многих лет
Личный опыт каждого из нас говорит, что индивидуальные воспоминания и навыки способны поддерживаться на протяжении очень длительно времени, иногда всю жизнь. Для героя М. Пруста вновь испытанный вкус бисквитного пирожного повлек за собой богатейшую череду юношеских воспоминаний, развернувшихся в многотомную эпопею «В поисках утраченного времени». Нередко воспоминания практи- чески не используются на протяжении многих лет и, тем не менее, при определенных условиях извлекаются из памяти. В клинической литературе описан случай, когда человек после потери сознания, связанной с ушибом мозга, заговорил на своем родном валлийском наречии, на котором он не говорил в течение 30 лет и, казалось бы, совершенно забыл его.
Однако для извлечения давней памяти не обязательно нужны необычные условия. Исследования здоровых людей, которые в школе обу- чались испанскому языку, но никогда потом им не пользовались, показали, что около 50% этого знания продолжало быть доступным и после 50-летнего перерыва. В некоторых случаях такая долговременная память может сохранять потрясающую точность. Нейропсихолог А. Р. Лурия, исследовавший память мнемониста Ш., описывает, как тот с точностью воспроизводил длинные ряды из 50–70 чисел или несвязанных слов спустя 15–16 лет после того, как он их единожды видел.
Но длительная и надежная память не является способностью лишь неординарных людей. При исследованиях зрительной памяти у обыч-
247
ных испытуемых было установлено, что они без труда узнавали около 90% фотографий своих одноклассников, с которыми они ни разу не встречались 35 лет после окончания школы. Каким же образом наша нервная система способна поддерживать память на протяжении столь длительного времени?
9.2. Поиски механизмов долговременной памяти направляются идеей энграммы — следа памяти в нервной системе
Более трех с половиной веков назад в «Страстях души» Р. Декарт предпринял первую попытку объяснить, как память образуется, хранится и извлекается мозгом: «Когда в душе возникает желание что-то вспомнить, оно заставляет [эпифизную] железу, попеременно наклоняющуюся в разные стороны, направлять «животные духи» в различ- ные части мозга до тех пор, пока они не встретят следов, оставленных тем предметом, который душа хочет вспомнить. Эти следы — не что иное, как приобретенное свойство пор мозга, через которые раньше проходили «духи», вызванные этим предметом, открываться с большей готовностью «духам», повторно приходящим к ним. Таким образом, «духи», встречая эти поры, входят в них легче, чем в другие, и вызывают особое движение в железе, передающее душе этот предмет и указывающее ей на то, что он и есть тот самый, который она хотела вспомнить».
Декартовская идея упражнения «пор» с помощью «животных духов», на двести лет предварившая открытие нервного возбуждения, нейронов и синаптических контактов, стала путеводной нитью для поисков связи памяти и мозга. Через полтора века Ж. Б. Ламарк повторял ее в «Философии зоологии» уже следующим образом: «При всяком действии, вызванном нервным флюидом, происходит перемещение этого флюида. Когда это действие многократно повторяется, то несомненно, что флюид, обусловливающий его, прокладывает себе путь, прохождение которого делается с течением времени для него тем более легким, чем чаще он им пользуется и чем сильнее выражена склонность флюида следовать именно по этому привыч- ному пути, а не по какому-либо иному, по которому он не столь часто движется».
В 1893 г., уже после создания клеточной теории и начала микроскопических исследований строения нервной системы, итальянский нейрогистолог Э.Танци (E.Tanzi, 1893) писал:
248
«Вероятно, каждое отображение чего-либо в мозгу сразу же вызывает функциональную гипертрофию протоплазматических выростов и аксонов; молекулярные вибрации усиливаются и распространяются, изменяя форму дендритов; таким образом, при благоприятных обстоятельствах возникают, а потом и сохраняются новые выросты
èответвления… каждый последующий эпизод одного и того же акта сознания в форме воспоминания, фантазии или единичного повторения усиливает мнемонический потенциал, поскольку ведет к формированию новых впечатлений, которые заменяют или дополняют существовавшие ранее… При этом на смену первичным кратковременным связям между нейронами в конце концов приходят стабильные функциональные объединения клеток… Этот прогрессирующий процесс функциональной гипертрофии приводит к более или менее стойкому увеличению разветвленных отростков, соединяющих нервные элементы».
Это предположение вместе с догадкой великого испанского нейроанатома C. Рамона-и-Кахала (Ramon y Cajal, 1893) о том, что формирование опыта вовлекает такие же клеточные механизмы, как
èразвитие нервной системы, составили основу грандиозной редукционистской программы исследований памяти в 20 веке. Ее руководящей идеей стал поиск в нервной системе следа памяти, или «энграммы». Понятие «энграмма» ввел немецкий биолог Р. Семон (R. Semon), опубликовавший в 1904 г. монографию «Die Mneme», в которой он пытался соединить биологический анализ наследственности c психологическим и физиологическим анализом памяти. Эти идеи не были оценены современниками, однако сам термин «энграмма» получил широкую популярность, в значительной степени благодаря статье знаменитого американского нейропсихолога К. Лешли (K. Lashley, 1950). В работе, называвшейся «В поисках энграммы» и подводившей итог более чем 30 годам труда, Лешли с горечью признавался: «Обозревая доказательства локализации следа памяти, я иногда ощущаю, что обучение просто невозможно. Слишком сложно представить себе механизм, который отвечал бы требованиям к этому процессу». Однако исследования механизмов памяти во второй половине 20 века позволили путем цепи детерминисти- ческих шагов перекинуть мост от «свойства человеческой души», как определяли память психологические руководства начала века, к тончайшим молекулярным процессам в нервной системе. Ниже мы рассмотрим основные звенья этой связи.
249