

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfПри активации медиаторами соответствующих им рецепторов постсинапс обеспечивает многообразные физиологические эффекты, определяемые особенностями функционирования рецепторов.
7.3. Диффузная передача сигнала
Медиаторы активируют соответствующие им рецепторы через синапсы или внесинаптическим способом. В отличие от синаптической, внесинаптическая передача в большей степени зависит от скорости диффузии медиатора в межклеточном пространстве. Внесинаптическая диффузная передача может изменять состояние большой группы нейронов, обладающих внесинаптическими рецепторами, чувствительными к диффундирующему медиатору. Внесинаптические рецепторы для некоторых медиаторов обнаружены на мембранах различных частей нейрона. Диффузное распространение медиатора из синаптической щели на некоторое расстояние называют спиловером (англ., spillover, spill — разливаться, over — перелив) (рис. 7.8).
Ðèñ. 7.8. Внесинаптическая передача. Выделившийся в синаптической щели медиатор растекается за пределы синаптического контакта и активирует внесинаптические рецепторы. Внесинаптические рецепторы могут активироваться медиатором, диффундирующим из соседнего по отношению к данному нейрону синапса. Также показана утечка медиатора — спиловер (по: Семьянов, 2003).
7.3.1. Источники диффузного сигнала
Описано несколько источников сигнала диффузной внесинапти- ческой передачи (рис. 7.9). Высвобождение нейропередатчика во внеклеточное пространство может происходить за счет обратно направ-
210
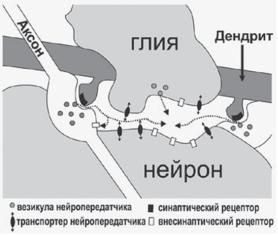
ленного захвата глутамата и ГАМК. Существуют специальные транспортные системы, которые обеспечивают захват медиаторов из внеклеточного пространства. Найдено несколько транспортеров, которые переносят глутамат через мембрану глиальных клеток и нейронов.
Ðèñ. 7.9. Источники диффузного внесинаптического медиатора: (1) обратно направленный транспорт медиатора из нейрона во внеклеточное пространство,
(2) выделение медиатора из глиальной клетки и (3) спиловер медиатора от близкорасположенных синапсов. Пунктирными стрелками обозначено направление диффузии медиатора. Внесинаптические рецепторы, расположенные на пресинаптических терминалях, выполняют функцию ауторецепторов.
Работа транспортеров глутамата основана на градиенте ионов Na+, K+ и самого медиатора. Процесс транспорта является электрогенным, поскольку внутрь клетки переносится положительный заряд. При гиперполяризации захват глютамата из внеклеточного пространства усиливается, а при деполяризации, напротив, подавляется.
Транспорт глутамата является симметричным процессом и может происходить в обоих направлениях. В зависимости от электрохимического градиента ионов Na+, K+, самого глутамата и потенциала мембраны глютамат будет либо поглощаться внутрь клетки, либо высвобождаться во внеклеточное пространство. Таким образом, обратно направленный захват глутамата транспортерами является источником внеклеточной концентрации этого медиатора.
Аналогичные транспортные системы найдены также и для ГАМК. Транспортеры ГАМК могут транспортировать ее через мембрану в обоих
211
направлениях. Изменение направления транспорта может происходить при изменении градиента Na+, деполяризации мембраны и подавлении активности ГАМК-трансаминазы.
Кроме самих нейронов, глиальные клетки также обладают механизмом высвобождения медиаторов посредством экзоцитоза. Такой экзоцитоз является Са2+-зависимым процессом и может запускаться при активации глиальных глутаматных рецепторов.
Еще одним источником внеклеточных глутамата и ГАМК является диффузия этих медиаторов из синаптической щели (спиловер).
7.3.2. Приемники диффузного сигнала
Приемниками в диффузной медиаторной передаче являются внесинаптические рецепторы, чувствительные к данному медиатору (рис. 7.10). Собственно синаптические рецепторы, расположенные на постсинаптической мембране в месте экзоцитоза медиатора, также способны активироваться диффузным глутаматом или ГАМК. Однако из-за относительно низкой аффинности синаптические рецепторы незначительно активируются низкими концентрациями диффузно выделяемого медиатора.
Основным механизмом удержания высокомолекулярного комплекса рецептора в синапсе является его взаимодействие с белками цитоскелета. Таких адаптерных белков, удерживающих рецепторы в синапсе, обнаружено достаточно много. Взаимодействие рецепторов с адаптерными белками, которое приводит к изменению числа рецепторов, регулируется рядом факторов. Таким образом, метаболические реакции в клетке способны менять синаптическую эффективность, изменяя число синаптических рецепторов за счет их перемещения во внесинаптический пул рецепторов и обратно. С другой стороны, изменение числа рецепторов во внесинаптическом пуле может оказывать влияние на эффективность диффузной нейропередачи.
Как и синаптические, внесинаптические рецепторы управляют либо ионным током, либо активируют метаболические внутриклеточные каскады. Естественно, ключевым фактором является тип нейрона, в котором возникает ответ на диффузный сигнал. Так, повышение возбудимости пирамидных клеток гиппокампа будет приводить к общему повышению возбудимости всей структуры в отличие от повышения возбудимости иннервирующих их тормозных интернейронов. В последнем случае будет происходить повышение торможения пирамидных клеток.
212

Ðèñ. 7.10. Синаптические и внесинаптические рецепторы. Внесинаптические рецепторы находятся за пределами синаптического уплотнения, как на постсинаптической клетке, так и пресинаптической терминали. Рецепторы, взаимодействующие с адапторными белками (РАБ), способны образовывать кластеры напротив места синаптического экзоцитоза. Таким образом адапторные белки связывают данные рецепторы с белками цитоскелета. При разрыве такой связи РАБ способны покидать синапс и перемещаться во внесинаптическую мембрану. Рецепторы, не взаимодействующие с адапторными белками (РНАБ), находятся преимущественно вне синапсов.
Помимо типа нейрона, эффект активации внесинаптических рецепторов зависит от участка нейрона, на котором они расположены. Активация пресинаптических метаботропных рецепторов влияет на высвобождение медиатора. С другой стороны, активация аксональных и со- мато-дендритных ионотропных внесинаптических рецепторов приводит к деполяризации или гиперполяризации мембраны. Такие изменения потенциала мембраны влияют на порог генерации потенциалов действия, на паттерн импульсных разрядов и на кинетику синаптических токов.
Таким образом, диффузный глутамат и ГАМК обеспечивают различные типы сигнализации в нейронных сетях. С одной стороны, изменение возбудимости нейрона при активации его ионотропных внесинаптических рецепторов диффузным медиатором аналогично возбуждающей или тормозной синаптической передаче (сигнальная функция). С другой стороны, синаптическая и диффузная передачи обладают сложным механизмом взаимодействия друг с другом. Спи-
213
ловер медиатора при синаптическом событии является источником диффузного сигнала. Активация пресинаптических рецепторов диффузным медиатором изменяет эффективность синаптической переда- чи (модуляторная функция).
Помимо глутамата и ГАМК, некоторые медиаторы (моноамины, ацетилхолин, пурины и др.) в ЦНС выполняют модуляторную функцию. В этом случае медиаторы не вовлекаются непосредственно в процессы синаптического возбуждения или торможения, а оказывают на них модулирующее влияние. Например, сбалансированная работа синаптического возбуждения/торможения и нейромодуляторных систем лежит в основе передачи и обработки информации в гиппокампе, а также генерации его ритмов.
7.4. Пресинаптический кальций: функция, регистрация и утилизация. Постсинаптические потенциалы и токи.
«Быстрые» и «медленные» синапсы
Скорость передачи сигнала в синапсе является одним из основных параметров при эволюционном выборе механизма межнейронного соединения. Основное преимущество электрического способа передачи сигнала между клетками является скорость проведения сигнала, то есть отсутствие синаптической задержки. Но и для химических синапсов скорость проведения часто определяет эволюционный выбор синапса данного типа (например, «чашечки Хелда»). Длительность синаптической задержки в химических синапсах в отличие от электрических может меняться, то есть является одним из факторов синаптической пластичности. В химических синапсах величина синаптической задержки определяется несколькими при- чинами:
—параметрами пресинапса (тип медиатора, тип и количество ионных каналов, вероятность выброса кванта медиатора (из 1 везикулы), число и расположение активных зон и число готовых к выбросу медиатора везикул, состояние пресинаптических «хелаторов»: кальциевых депо
èò. ä.);
—параметрами синаптической щели (ширина, ионный состав, ферменты, утилизирующие медиатор, и ферменты обратного захвата медиатора);
—параметрами постсинапса: тип рецепторов (ионо- и метаботропные); число и расположение рецепторов (постсинатические уплотнения).
214

7.5. Возбуждающие и тормозные синапсы
Одним из оснований для разделения медиаторов на типы долгое время служил эффект, который они вызывают (рис. 7.11). Говорили о возбуждающих и тормозных медиаторах и, соответственно, синапсах. Но это неправильно (вернее, правильно не абсолютно). Один и тот же медиатор может выполнять и возбуждающую, и тормозную функцию.
À |
Á |
Ðèñ. 7.11. Классификация синапсов по результату действия: возбуждающие и тормозные синапсы. Возбуждение и торможение, по Дж. Экклсу (1966). Рисунок нейрона, от тела которого отходят дендриты и аксон и к которому подходят аксонные терминали других нейронов. Оригинальные рисунки из: Sir John Carew Eccles — Nobel Lecture, December 11, 1963: The Ionic Mechanism of Postsynaptic Inhibition (по: Экклс, 1966).
7.6.Эргичность. Медиаторы: нейротрансмиттеры
èнейромодуляторы, комедиаторы
Медиаторная специфичность химического синапса (эргичность) является одной из основных его характеристик. Необходима идентификация вещества, передающего сигналы в синаптическом соединении. Наиболее часто используемые названия для таких химических посредников — это медиатор (от латинского mediator — посредник, или нейромедиатор) и трансмиттер (нейротрансмиттер).
В настоящее время определена медиаторная функция для десятков химических веществ (сотен — с вариациями для медиаторов-нейро- пептидов). Почему нейроны различаются по эргичности? Этот вопрос
215
требует специального, эволюционного, рассмотрения (Сахаров, 1990). Эволюционный подход дал однозначный ответ: «Химическое разнообразие синаптических механизмов, представленных в нашем мозге, обусловено...условиями возникновения нервной системы у примитивных многоклеточных животных.» (Сахаров, 1974, с. 164).
7.7. Классификация синапсов по источнику и мишени, сложные синапсы
Аксо-аксонный синапс
Дендро-дендритный синапс
Последовательные синапсы |
Синаптические гломерулы |
|
(аксо-аксо-дендритные) |
||
|
Глиальная
обертка
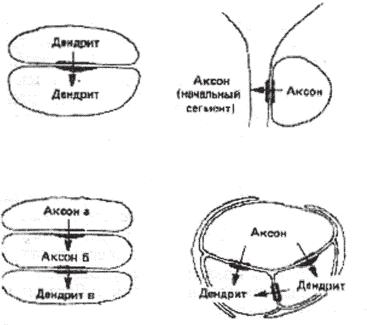
Ðèñ. 7.12. Классификация синапсов по источнику и мишени. Синапсы, различающиеся по типу пресинаптического волокна, по постсинаптической мишени и по сложности. Обнаружены все варианты контактов: аксо-денд- ритные (А-Д), — аксо-аксональные (А-А), аксо-соматические (А-С), дендродендритные (Д-Д), соматические, дендро-аксональные (Д-А) и их сочетания, сложные синапсы, синаптические гломерулы (А-А-Д, А-Д-Д) (Шеперд, 1987).
Распространенный способ классификации синапсов — по обозна- чению структуры пре- и постсинаптических нейронов, формирующих синапс (Шеперд, 1987). Термины аксо-дендритный, аксо-соматический
216
и аксо-аксональный (с соответствующей аббревиатурой) прочно вошли в литературу по нейронаукам. Показана возможность формирования контактов всех возможных типов. Так, термины дендро-дендритный синапс или дендро-аксональный синапс сегодня не удивляют, хотя вызывают споры. Например, возникает вопрос о «дендритных спайках». Возможность генерации ПД в дендритах экспериментально показана, но пока эти данные считаются исключением из общего правила об электротоническом проведении потенциалов в дендритах. Экспериментально показано и существование устойчивых синаптических структур, образованных более чем двумя клетками. Например, есть данные о «последовательных» аксо-аксо-дендритных и аксо-дендро-дендрит- ных синапсах (рис. 7.12). Боковые (латеральные) и возвратные (обратные) связи между нейронами являются для НС скорее правилом, чем исключением.
Споры возникают из-за кажущегося нарушения принципа «динамической поляризации» нейрона. Но с нарастающим потоком данных о распространенности в НС «обратных связей» приходится считаться. Другое дело, что большинство таких связей реализуется с помощью химических, но не синаптических механизмов взаимодействия нейронов (объемная передача, параэндокринное взаимодействие. Роуз, 1995, Сахаров, 1990). Об этом говорят и данные об общности рецепторых механизмов при синаптическом и несинаптическом способах взаимодействия нейронов.
7.8. Обратные посредники
Рецепторы нейромедиаторов находятся не только на постсинапти- ческой мембране, но и в области пресинаптической терминали. Для некоторых рецепторов (например, H3 — рецепторы гистамина, HA) пресинаптическая авторегуляция является типичным способом функционирования («аутосинапсы»). По паракринному способу передачи сигнала (объемная передача) функционируют другие, «обратные посредники», передающие сигнал через специализированные рецепторы (нейротрофины, нейротропины и их TRK-R). Обратные посредники (рис. 7.13) рассматриваются как обязательное звено передачи сигналов в пластичных синапсах («подкрепление» на клеточном уровне). Не все обратные посредники передают сигнал, активируя рецепторы и влияя на эффективность синаптической передачи через другие механизмы (инозитол-фосфат, IP3, NO и Ca2+).
217
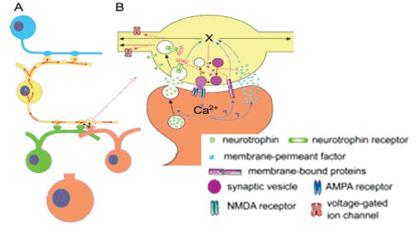
Ðèñ. 7.13. Обратные посредники.
7.9. Идентифицируемые нейроны и синапсы
Многообразие типов нервных клеток в мозге, в том числе функциональное, предполагает разнообразие синаптических контактов между ними. И это действительно так. Поэтому один из наиболее распространенных способов классификации синапсов — это определение через название контактирующих клеток. Самое общее определение — по функции, например, нервно-мышечный синапс, сенсо-моторный синапс. Более детальное — через название афферентного волокна, формирующего синапс, и название постсинаптического нейрона. Например, синапс между мшистыми волокнами и клеткой Пуркинье мозжечка или синапс между лазающими волокнами и пирамидными нейронами поля СА3 гиппокампа. В последнее время все большее распространение находит обозначение «идентифицированные синапсы млекопитающих». Началом этому было изучение синаптических контактов между индивидуально идентифицированными нервными клетками беспозвоночных животных, например, у моллюсков межнейронные синапсы определялись по номенклатуре идентифицированных нейронов (Кэндел, 1980).
Возможность сочетания в одном исследовании морфологических, электрофизиологических, молекулярно-генетических и т.п. методов в сочетании с развитием компьютерной техники позволило значительно продвинуться в понимании функционирования мозга на клеточном и
218
молекулярном уровнях. А возможность идентификации нейронов, то есть возможность совместного использования количественных данных, полученных в разных лабораториях, объединяемых моделями, и проверяемых с помощью их компьютерной реализации, позволяет говорить о «нанонейронике» (Соколов, 2006). Новый пик робототехники в начале нового тысячелетия является во многом отражением достижений нейронаук на этом, нанонейронном, уровне.
Механизмы функционирования синаптических контактов на нанонейронном уровне можно представить на конкретных примерах идентифицированных синапсов беспозвоночных животных (на «простых нервных системах») и млекопитающих.
К идентифицированным синапсам относят:
—Идентифицированные синапсы моллюсков (аплизия и виноградная улитка).
—Идентифицированные синапсы позвоночных: нервно-мышечные соединения, хромаффинные клетки, сетчатка, центральные синапсы млекопитающих.
Идентифицированными называют синапсы между идентифицированными нейронами (Соколов, 2003). Под идентифицируемым нейроном понимают такую нервную клетку, которую можно надежно определить по ее локализации в НС, размеру, типу электрической активности, реакции на стимулы и по другим параметрам от эксперимента к эксперименту. Такому требованию прежде всего соответствуют гигантские нейроны виноградной улитки, которые расположены на поверхности ганглиев, доступны наблюдению и удобны для внутриклеточной регистрации.
Существует и более широкое значение термина «идентифицируемый нейрон». Это нейрон определенного морфологического, физиологического и функционального типа в определенной структуре мозга. Такие нейроны можно идентифицировать на препаратах нервной системы высших животных, включая млекопитающих, хотя об индивидуальной идентификации, как в нервной системе моллюсков, говорить не приходится. Тем не менее, понятие об «идентифицируемым синапсе» для нервной системы беспозвоночных и млекопитающих полностью совпадает. Для идентификации синапса основным требованием является возможность одновременной внутриклеточной регистрации активности пре- и постсинаптического нейронов. Такую одновременную регистрацию чаще называют «парной». Главное требование для парной регистрации — идентификация пре- и постсинаптического нейронов.
219