

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfВыделившийся из сенсорного нейрона медиатор глутамат вызывает ВПСП в мотонейроне (рис. 8.2, слева), который, в свою очередь, активируя мышцы жабры, вызывает оборонительную реакцию — втягивание жабры в полость тела.
Ðèñ. 8.1. Нейронная сеть, обеспечивающая оборонительную поведенческую реакцию аплизии. Пунктиром выделен фрагмент, включающий мотонейрон и аксонные терминали сенсорного нейрона и фасилитирующего интернейрона.
Усиление такой простой реакции получают, добавляя возбуждение другой части тела, например, хвоста. Механическое или электрическое раздражение хвоста возбуждает сенсорный нейрон, который, в свою очередь, активирует фасилитирующий интернейрон (рис. 8.1, сверху). Аксонные терминали интернейрона оканчиваются в том числе и на пресинаптических терминалях сенсорного нейрона (от сифона), иннервирующего мотонейрон. Выделяющийся из фасилитирующего интернейрона серотонин оказывает модулирующее действие на синаптическую передачу от сенсорного нейрона, иннервирующего кожу сифона, к моторному нейрону. Модулирующее влияние серотонина приводит к
230
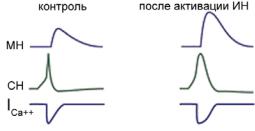
увеличению выброса медиатора из сенсорного нейрона, что вызывает увеличение амплитуды ВПСП в мотонейроне (рис. 8.2, справа) и сенситизацию оборонительной реакции моллюска.
Ðèñ. 8.2. ВПСП мотонейрона (МН) и сенсорного нейрона (СН) в ответ на стимуляцию только кожи сифона (слева) и при дополнительной стимуляции хвоста (справа). ICa++ — Ñà2+-ток, входящий в аксонную терминаль сенсорного нейрона. Объяснения в тексте.
Роль серотонина в механизме сенситизации оборонительной реакции моллюска подтверждают эксперименты с прямой аппликацией серотонина на пресинаптическую мембрану сенсорного нейрона. Прямое действие серотонина вызывает усиление двигательного ответа так же, как и механическая или электрическая стимуляция хвоста.
8.2.2. Молекулярные механизмы кратковременной фасилитации
Взаимодействуя с мембранными рецепторами пресинаптической мембраны сенсорного нейрона, серотонин через 5НТ4-рецепторы инициирует каскады синтеза цАМФ, активируя через Gs-белки аденилатциклазу (рис. 8.3). цАМФ активирует цАМФ-зависимую протеинкиназу А, которая фосфорилирует потенциал-зависимые Кs+-каналы. Фосфорилирование этих каналов уменьшает К+-проводимость (рис. 8.3, путь 1), что приводит к удлинению деполяризации и увеличению продолжительности Са2+-токов (рис. 8.2), текущих через каналы N-типа.
Кроме того, протеинкиназа А фосфорилирует потенциал-зависи- мые Са2+-каналы L-типа, устраняя их инактивацию, что приводит к еще большему притоку Са2+ в пресинаптическую терминаль (рис. 8.3, путь 3).
Избыточный Са2+ в комплексе с белком калмодулином активирует Са2+/калмодулин-зависимую протеинкиназу II (СаМК II) (не показано). Эта протеинкиназа фосфорилирует белок везикул синапсин, ос-
231
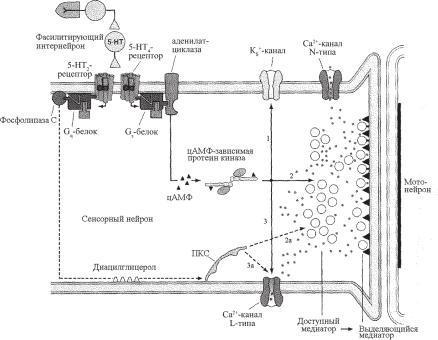
лабляя его связь с актиновыми нитями цитоскелета, которые удерживают везикулу в цитоплазме. В результате увеличивается мобилизация везикул из их общего пула в активной зоне, и они в большем количе- стве транспортируются к месту высвобождения медиатора (рис. 8.3, путь 2). Другой белок везикулы синаптотагмин является сенсором Са2+ и запускает стыковку и слияние мембраны везикулы с пресинаптической мембраной (не показано). Избыток Са2+ положительно влияет на адгезию везикул с пресинаптической мембраной.
Ðèñ. 8.3. Схема внутриклеточных каскадов механизма кратковременной сенситизации оборонительной реакции моллюска. Объяснения в тексте.
Другой серотониновый рецептор 5НТ2–типа активирует Gq-белки, которые запускают каскад мембранных липидов. Активированная Gq- белками фосфолипаза С расщепляет мембранный липид ФИФ2 íà ÈÔ3 и ДАГ. Последний активирует протенкиназу С (ПКС), мишенями которой являются Са2+-каналы L-типа, а также процессы мобилизации везикул и их адгезии с преиснаптической мембраной (рис. 8.3, пути 2а и 3а).
232
8.2.3. Механизмы долговременной фасилитации сенсомоторных связей в нервной системе моллюсков
При определенном числе повторений гетеросинаптической стимуляции концентрация цАМФ под действием серотонина достигает высоких значений, достаточных для формирования долговременных изменений синаптической эффективности. Долговременная модификация синапти- ческой передачи определяется функциональными и структурными изменениями, связанными с активацией экспрессии некоторых генов.
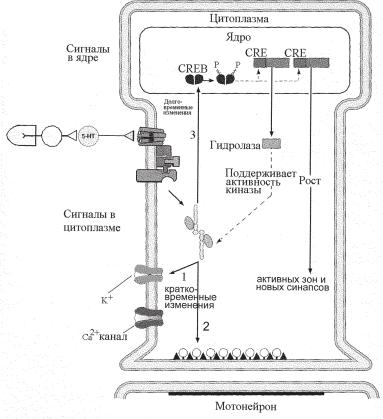
Ðèñ. 8.4. Схема внутриклеточных каскадов механизма долговременной сенситизации оборонительной реакции моллюска. Пути 1 и 2 отражают каскады, обеспечивающие кратковременную сенситизацию (см. рис. 8.3). Объяснения в тексте.
233
Упрощенная схема механизма долговременной сенситизации представлена на рисунке 8.4. Серотонин, выделяющийся из фасилитирующего интернейрона, вызывает увеличение синтеза цАМФ. Ключевую роль в долговременной фасилитации играет цАМФ-зависимая протеинкиназа А, которая активирует экспрессию генов путем фосфорилирования факторов транскрипции (рис. 8.4, путь 3) — регуляторного белка ApCREB1 (Aplysia CRE-binding protein), идентифицированного у аплизии (на рис. 8.4 обозначен как CREB). Кроме этого белка, у аплизии выделен другой белок ApCREB2 (не показан) с противоположной функцией, который блокирует транскрипцию цАМФ-активируемых генов. Долговременная фасилитация достигается не только активацией факторов транскрипции, но и ингибированием репрессоров транскрипции. Так, серотонин активирует митоген-зависимую протеинкиназу (не показана), которая ингибирует репрессор транскрипции ApCREB2 путем его фосфорилирования.
Факторы транскрипции усиливают синтез, по крайней мере, десяти различных белков. Эти белки по принципу положительной обратной связи усиливают выделение медиатора из сенсорного нейрона. Один из этих белков (гидролаза, рис. 8.4) вызывает деградацию регуляторных субъединиц протеинкиназы А, что приводит к усилению функции каталитических субъединиц. Поддерживаемая активность каталитических субъединиц протеинкиназы А приводит к эффектам, обеспечивающим кратковременные изменения синаптической передачи (рис. 8.4, пути 1 и 2). Кроме того, активная протеинкиназа А поддерживает фосфорилирование АрCREB1, обеспечивая положительную обратную связь. Еще один белок способствует росту активных зон пресинаптической мембраны и развитию новых синаптических контактов.
8.2.4.Пресинаптическая пластичность командных нейронов улитки
Фокусом ассоциативной пластичности в мозге является изменение синаптических весов. Моллюски помогают понять синаптические механизмы формирования памяти (Kandel, 2000). Возможность идентифицировать нейроны и синапсы делает моллюсков излюбленной экспериментальной моделью нейробиологов (Dullock, 2000). Изменение чувствительности пресинаптических рецепторов в зависимости от постсинаптического выхода может представлять механизм ассоциативной селективности. Это предположение основано на данных эксперимен-
234
тального исследования идентифицированных синапсов наземной улитки (Helix lucorum). Гипотеза «пресинаптического ожидания» состоит в том, что пресинаптически активные терминали меняют свою чувствительность к таким обратным посредникам, как нейротрофические факторы, выполняющим функцию «подкрепления».
Пресинаптические изменения, зависящие от постсинаптической активности, были зарегистрированы в «простых нервных системах» и в мозге позвоночных животных. Парная регистрация, одновременная внутриклеточная регистрация синаптически связанных идентифицированных нейронов виноградной улитки, показала, что даже два пресинаптических потенциала действия (ПД) вызывают увеличение амплитуды постсинатического ответа (унитарного возбуждающего постсинаптического потенциала, унитарного ВПСП, uEPSP) на третий пресинаптический ПД, используемый в качестве теста. Квантовый анализ унитарных ВПСП, вызванных в командных нейронах пассивно-оборонительного поведения пресинаптическими ПД в париетальных сенсорных нейронах, показал, что у улитки кратковременная пластичность, фасилитация (от англ. facilitate — облегчать, помогать, способствовать — облегчение, повышение эффективности проведения сигнала), огментация (повышение амплитуды потенциала), посттетаническая потенциация и зависящая от ча- стоты депрессия имеют пресинаптическое происхождение. Синаптические изменения (позитивная или негативная пластичность) зависят от паттерна пресинаптических ПД. Двухкомпонентная структура суммарных ВПСП (сВПСП), регистрируемых в командных нейронах улитки в ответ на одиночные сенсорные стимулы, является результатом немедленной (immediate) пластичности. Баланс между позитивной и негативной пластичностью проявляется в стабильности амплитуды ВПСП в ответ на ритмическую стимуляцию определенной частоты.
Гипотеза «пресинаптического ожидания» подтверждается результатами исследований на других экспериментальных моделях, включая нервно-мышечные соединения лягушки и центральные синапсы млекопитающих.
8.3. Постсинаптическая пластичность
Феномен ДВП заключается в продолжительном усилении постсинаптического ответа после тетанической активации (высокочастотной серией импульсов, 100 Гц, 50–100 стимулов) в ответ на стимуляцию входа, который до тетанизации вызывал слабый ответ. ДВП вызывается тремя способами: гомосинаптическая ДВП возникает в активи-
235
рованном синапсе как результат его собственной активации; гетеросинаптическая ДВП возникает в синапсе в результате активации другого синапса; разновидность гетеросинаптической ассоциативная ДВП возникает в синапсе в результате его одновременной активации с другим синапсом. Феномен ДВП впервые был обнаружен в пирамидных клетках гиппокампа и интенсивно исследуется как клеточный аналог памяти и обучения.
8.3.1.Гомосинаптическая ДВП
âпирамидных клетках гиппокампа
ДВП была впервые описана Т. Блисс и Т. Ломо в глутаматэргических синапсах гиппокампа (рис. 8.5). Было показано, что короткая высокочастотная стимуляция перфорантного пути (синаптического входа к зернистым клеткам зубчатой фасции) приводит к увеличению амплитуды суммарного постсинаптического ответа этих клеток, которое сохраняется в течение десятков минут и часов, а при специальных условиях — дней, недель и даже месяцев, то есть интервалов времени, сопоставимых с поведенческой памятью.
8.3.2.Ассоциативная ДВП
âпереживающих срезах гиппокампа
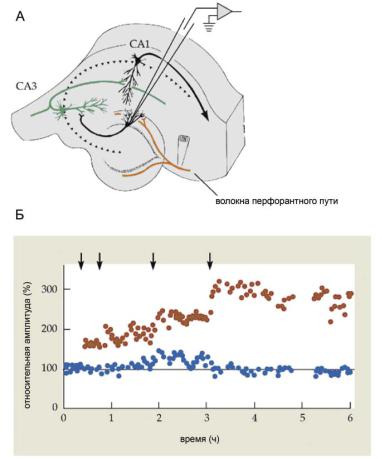
Ассоциативная ДВП в переживающих срезах гиппокампа представляет собой результат ассоциации возбуждения двух афферентных путей. В экспериментах пирамидные нейроны поля СА1 стимулируют через два пути (рис. 8.6, А). Первый путь — это «слабая» стимуляция коммисуральных входов, которая вызывает слабый ПСП. Второй путь — «сильная» стимуляция коллатералей Шаффера (входы от нейронов CA3), которая вызывает существенно больший ПСП.
Тетанизация коротким залпом импульсов (100 Гц в течение 1 секунды, повторенный через 5 с) через «сильный» вход привела к усилению ответов, активируемых через этот же вход (гомосинаптическая ДВП, аналогичная приведенной на рис. 8.5). Тетанизация через «слабый» вход (рис. 8.6, Б, второй ответ) так же, как и тетанизация через «сильный» вход (рис. 8.6, Б, третий ответ), не оказали влияния на величину ответа, вызываемого через «слабый» вход. Однако после совместной стимуляции «сильного» и «слабого» входов наблюдалось существенное увеличе- ние величины ответа в ответ на стимуляцию «слабого» входа (рис. 8.6, Б, четвертый ответ), продолжающееся десятки минут (рис. 8.6, В). Этот феномен в современной нейробиологии рассматривается как клеточный
236
аналог ассоциативного обучения (подобного синапсу Хэбба, когда стимуляция «сильного» входа ассоциируется с безусловным, а стимуляция «слабого» входа — с условным стимулами (рис. 8.7).
Ðèñ 8.5. Гомосинаптическая ДВП в гиппокампе анестезированного кролика. (А) Синаптические ответы на стимуляцию перфорантного пути регистрировали в гранулярных клетках зубчатой фасции. (Б) Короткие тетанусы (15 стим./ с в течение 10 с) наносятся в отмеченные стрелками моменты. После каждого тетануса отмечается относительное увеличение синаптического ответа (красные точки). В контрольной серии (без тетанусов) изменений синаптического ответа не наблюдается (синие точки).
237
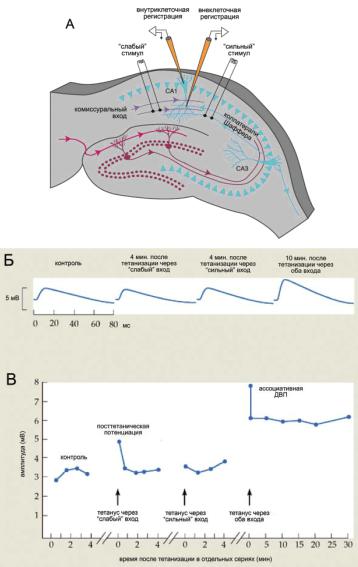
Ðèñ. 8.6. Ассоциативная ДВП в срезах гиппокампа. (А) Схема эксперимента; (Б) усредненные ответы; (В) суммированные результаты. Объяснения в тексте.
238
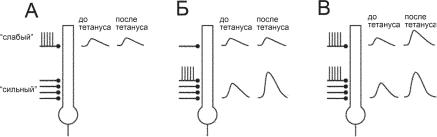
Ðèñ. 8.7. Модель классического условного рефлекса (ДВП) на нейронах гиппокампа. Пирамидные нейроны поля СА1 активируются через «слабый» и «сильный» входы. «Слабый» и «сильный» входы в модели условного рефлекса соответствуют условному и безусловному стимулам.А. Тетаническая стимуляция только через «слабый» вход не вызывает усиления эффективности «слабого» синапса (ДВП) через этот путь.Б. Тетаническая стимуляция только через «сильный» вход вызывает усиление эффективности «слабого» синапса (ДВП) через этот путь.В. Тетаническая стимуляция через оба входа одновременно вызывает усиление эффективности «слабого» синапса (ДВП) через «слабый» путь.
8.3.3. Механизмы индукции ДВП
Возникновение ДВП связывают с дополнительным увеличением концентрации внутриклеточного Са2+ в постсинаптическом нейроне, вызванным совместной стимуляцией нейронов поля СА1 через «сильный» и «слабый» афферентные входы. Механизмом увеличения концентрации внутриклеточного Са2+ является активация глутаматных NMDA-рецепторов. NMDA-рецепторы характеризуются более сложными свойствами по сравнению с другими глутаматными рецепторами (каиновыми и AMPA), поскольку их проводимость зависит не только от медиатора, но и от уровня мембранного потенциала.
В условиях умеренной стимуляции глутаматэргического пресинаптического нейрона (стимуляция только через один вход в модели на срезах гиппокампа) в постсинаптическом нейроне генерируются непродолжительные ВПСП, вызванные Na+/K+-проводимостью только через активированные глутаматом AMPA и каиновые рецепторы. Если деполяризация незначительная, NMDA-рецепторы не активируются из-за Mg2+-блока, и регистрируется непродолжительный ВПСП (рис. 8.8, Б, контроль, В). При интенсивной стимуляции (стимуляция через два пути в модели на срезах гиппокампа, рис. 8.6) деполяризация достигает уровня
239