

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfÐèñ. 6.28. Воротные токи. (А) В условиях пониженной концентрации ионов Na+ в ответ на деполяризацию регистрируется малый по амплитуде выходящий ток, отражающий заряд мембранной емкости (а). В ответ на гиперполяризацию этот емкостной ток меньше, чем в ответ на деполяризацию такой же амплитуды (б). Разность этих токов (воротный ток, или асимметричный ток смещения) соответствует перемещению зарядов, например, повороту диполя (в). (Б) Воротный ток маскируется входящим Na+-током большой амплитуды. (В) Воротный ток отчетливо выделяется при ТТХ блокаде Na+-òîêà.
Из результатов по исследованию инактивации Na+-проводимости следует, что инактивация каналов является самостоятельным процессом. Последующие эксперименты с введением протеолитических (разрушающих белки) ферментов внутрь аксона показали замедление и дальнейшее исчезновение инактивации Na+-каналов. Использование этих ферментов с наружной стороны мембраны не вызывало устранения инактивации. Предположительно, эти ферменты разрушили часть белковой молекулы канала, отвечающей за инактивацию.
170
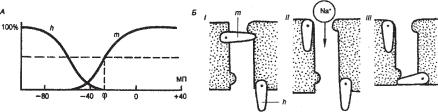
6.8.6. Основные состояния натриевых каналов (цикл А. Ходжкина)
На основании результатов по исследованию Na+-токов А. Ходжкин и А. Хаксли сформулировали представления об изменении состояния Na+-каналов во время возбуждения мембраны. В покое (мембрана не деполяризована) канал не пропускает Na+, поскольку закрыты m-воро- та (рис. 6.29, Б, I). При деполяризации открываются активационные m- ворота, канал активируется и пропускает Na+ (рис. 6.29, Б, II) При длительной деполяризации закрываются инактивационные h-ворота, расположенные у внутренней стороны мембраны, канал инактивируется и перестает пропускать Na+ (рис. 6.29, Б, III). Реполяризация мембраны до уровня ПП вновь приводит к открытию h-ворот и закрытию m- ворот, и цикл повторяется.
В цикле Ходжкина доли открытых m- è h-ворот реципрокно изменяются в зависимости от потенциала мембраны (рис. 6.29, А). При деполяризации примерно от -50 мВ и выше доля открытых m-ворот резко увеличивается, достигая уровня 50% при -30 мВ (отмечено пунктиром), однако в это время доля открытых h-ворот уменьшается до 0. Такая модель объясняет кратковременный характер входящего Na+-òîêà.
Ðèñ. 6.29. (А) Зависимость состояния m- è h-ворот потенциал-зависимых Na+-каналов от мембранного потенциала (МП). (Б) Схематическое изображение цикла А. Ходжкина. Объяснения в тексте.
6.8.7. Рефрактерный период
Во время генерации ПД в мембране происходят два процесса, из-за которых в нейроне или аксоне невозможно вызвать повторный ПД: (1) в начале нисходящей фазы ПД в мембране достигает максимума инактивация Na+-проводимости, и для ее устранения необходимо несколько миллисекунд; (2) активируются потенциал-зависимые К+-каналы, обес-
171
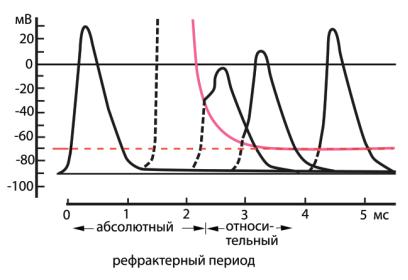
печивающие быструю реполяризацию мембраны. Эти два процесса приводят мембрану в состояние абсолютной рефрактерности, которое поддерживается в течение фазы спада ПД и характеризуется тем, что никакая деполяризация мембраны в этот период не способна запустить ПД (рис. 6.30). Период абсолютной рефрактерности сменяется периодом относительной рефрактерности, в ходе которого порог постепенно снижается до нормального значения. В этот период устраняется Na+- инактивация и потенциал-зависимая К+-проводимость. Абсолютный рефракторный период ограничивает максимальную частоту генерации ПД в нейронах и волокнах, препятствуя нарушению ионного баланса между клеткой и внеклеточной средой.
Эти результаты А. Ходжкина и А. Хаксли имеют фундаментальное значение для физиологии возбудимых мембран. Сделанные ими открытия представляют собой общий механизм регуляции состояния возбудимых мембран, способствующий восстановлению электрохимических градиентов, необходимых для нормального функционирования нейронов.
Ðèñ. 6.30. Периоды рефрактерности нерва во время и после возбуждения. В нерве млекопитающего вызывают ПД (слева), затем нерв стимулируют с разными интервалами. Сплошной красной линией показан пороговый уровень ПД, а черным пунктиром — деполяризация волокна стимулом до порогового уровня. В абсолютном рефрактерном периоде волокно невозбудимо, в относительном — порог его возбуждения превышает уровень в норме.
172

Ðèñ. 6.31. (А) Принципиальная схема метода локальной фиксации. (Б) Входящие токи, проходящие через одиночные Na+-каналы. Деполяризация локального участка мембраны клетки (а, Vp) приводит к кратковременному открыванию Na+-канала и к генерации входящего токов (б, Ip). Сумма таких токов сходна с Na+-макротоком мембраны аксона (в), отражающим срабатывание многих каналов в ответ на деполяризующий стимул.
173
6.8.8. Исследование ионных токов методом локальной фиксации
Для регистрации ионных токов, проходящих через отдельные каналы, была разработана методика локальной фиксации мембраны клетки. Для этой цели используют тонкие пипетки с диаметром около 1 мкм, которые тесно соприкасаются с клеточными мембранами (рис. 6.31, А). Высокое сопротивление такого контакта позволяет регистрировать слабые токи, проходящие через фрагмент мембраны, контактирующий с кончиком пипетки. Открытие и закрытие ионных каналов отражаются в виде прямоугольных токовых сигналов (рис. 6.31, Б). Свойства макротоков, впервые зарегистрированных в аконе кальмара, в дальнейшем были выявлены для токов отдельных каналов. Амплитуда токов, проходящих через отдельные Na+-каналы, так же, как амплитуда макротоков, зависела от мембранного потенциала. Кроме того, от мембранного потенциала зависела вероятность открытия каналов, отражая состояние m- è h-ворот. Деполяризация участка мембраны клетки приводит к кратковременному открыванию Na+-канала. Сумма таких токов сходна с Na+-макротоком мембраны аксона, отражающим срабатывание многих каналов в ответ на деполяризующий стимул.
6.8.9.Проницаемость Na+ через канал
Á.Хилле исследовал проницаемость Na+-канала для различных щелочных металлов и органических катионов. Он определил размер Na+-канала 0,3х0,5 нм и пришел к выводу, что Na+-связывающий уча- сток канала окружен кольцом атомов кислорода, входящих в состав гидроксильных групп мембранного белка.
Селективность канала зависит не только от размеров проникающих ионов, но и от их способности образовывать водородные связи. Б. Хил-
ле сравнивал проницаемости трех ионов: гидроксиламмония (H3N+-OH), гидразиния (H3N+-NH2) и метиламмония (H3N+-CH3). Радиусы гидроксильной, амино- и метильной групп примерно одинаковы и состав-
ляют 0.37–0.38 нм, однако проницаемость H3N+-OH оказалась в 100 раз выше, чем для H3N+-CH3. Такие различия объясняются тем, что метильные группы не образуют водородных связей. Длина водородной связи 0.08 нм, поэтому через канал проникают ионы размером не более 0.30 нм. На основании своих данных Б. Хилле заключил, что ионы проникают через канал не путем простой диффузии, а в результате последовательной дегидратации и связывания с компонентами канала.
174
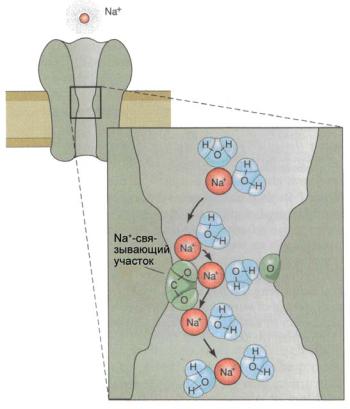
Ðèñ. 6.32. Прохождение иона Na+ через канал. В области Na+-связываю- щего участка гидратированный ион Na+ частично или полностью дегидратируется. Пройдя через селективный фильтр в наружном устье канала, ион вновь гидратируется.
В области селективного фильтра (Na+-связывающего участка) гидратированный ион Na+ частично или полностью дегидратируется из-за образования водородных связей гидратной оболочки с атомами кислорода входящих в состав гидроксильных групп (рис. 6.32). Избирательность каналов к ионам определяется стерическими (пространственными) свойствами селективного фильтра и составом химических групп, обеспечивающих электростатическое взаимодействие канала с данным типом иона. Пройдя через селективный фильтр в наружном устье канала, ион вновь гидратируется.
175
Канал является именно порой, и Na+ транспортируется через канал без какого-либо переносчика, так как переносчик способен транспортировать ионы со скоростью 104 ñ-1, а скорость диффузии через Na+-канал составляет 107–108 ñ-1.
6.8.10. Мультиионная природа токов в мембранах нейронов
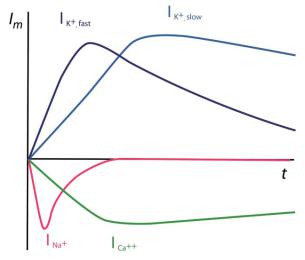
Исследование молекулярных механизмов возбудимости в мембранах нейронов стало возможным после разработки специальных методов, позволяющих исследовать мембраны таких миниатюрных объектов. В 1960-х г. под руководством П. Г. Костюка был разработан метод внутриклеточного диализа, позволяющий регистрировать токи, проходящие через мембраны нейронов. С использованием этого метода в мембранах нейронов беспозвоночных (виноградной улитки) были описаны потенциал-зависимые Na+- è Ê+-токи, аналогичные токам в мембранах гигантского аксона кальмара. Кроме того, используя специфи- ческие блокаторы, удалось идентифицировать еще один входящий ток, переносимый ионами Са2+, и дополнительный К+-ток, который, в отли- чие от «классического» аксонного тока, медленно активировался и медленно инактивировался (рис. 6.33). Позже эти токи были зарегистрированы и в мембранах морского моллюска Aplisia.
Ðèñ. 6.33. Схематическое изображение токов, зарегистрированных в мембранах нейронов виноградной улитки. Выделены два входящих (быстрый Na+-ток и медленный Ca++-ток) и два выходящих тока (быстрый IÊ+,fast и задержанный IÊ+,slow).
176

Ðèñ. 6.34. Схематическое изображение токов, зарегистрированных в мемб-
ранах нейронов позвоночных. Выделены три входящих (быстрый INa+,fast и медленный INa+,slow, медленный ICa++) и два выходящих тока (быстрый IÊ+,fast и задержанный IÊ+,slow). Медленный Na+-ток, в отличие от быстрого, оказался нечувствительным к ТТХ.
Кроме токов, обнаруженных в нейронах моллюсков, в мембранах более мелких нейронов позвоночных был зарегистрирован дополнительный Na+-ток, который не блокировался ТТХ и обнаруживался у 10–15% изученных нейронов (рис. 6.34). По кинетике этот ток напоминал медленный Ca2+-ток, и его можно было выделить, только заблокировав Са2+-проводимость ионами Со2+ èëè Cd2+.
Потенциал-зависимые катионные каналы могут быть локализованы не только в соме нейронов, но и в их дендритах. Так, в мембранах дендритов клеток Пуркинье были зарегистрированы потенциал-зави- симые Са2+-токи, которые обеспечивали генерацию ПД (рис. 6.35). Эти дендритные ПД достигают сомы нейрона, прерывая тоническую последовательность соматических Na+/K+ ПД. Соматические ПД не распространяются по всему дендритному дереву и лишь электротонически проникают на небольшие расстояния от сомы.
В дальнейшем были открыты и исследованы многочисленные по- тенциал-зависимые каналы для основных ионов, играющих определенную роль в механизмах активации клеточных мембран (рис. 6.36).
177
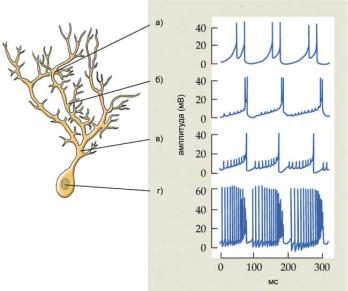
Ðèñ. 6.35. ПД в соме и дендритах клеток Пуркинье. В дендритах зарегистрированы ПД (а-в), обусловленные Са2+-проводимостью через потенциал-зави- симые Са2+-каналы. Эти дендритные ПД достигают сомы нейрона, прерывая тоническую последовательность соматических Na+/K+ ПД (г). Соматические ПД не распространяются по всему дендритному дереву и лишь электротони- чески проникают на небольшие расстояния от сомы (в).
Потенциал-зависимые каналы (и пропускаемые ими токи) разделяют по различным свойствам. Например, выделяют низкопороговые и высокопороговые токи, которые возникают, соответственно, при небольших и существенных колебаниях мембранного потенциала относительно
ПП. Другим критерием является временная динамика тока. В соответствии с этим критерием токи бывают фазические (англ., transient),
или быстро инактивирующиеся, и длительные — непрерывные
(sustained), устойчивые, постоянные (persistent), продолжительные (long lasting).
Na+-òîêè
Äâà Na+-тока широко распространены среди нейронов различных частей нервной системы. Эти токи различаются по порогу активации, степени инактивации и амплитуде.
178
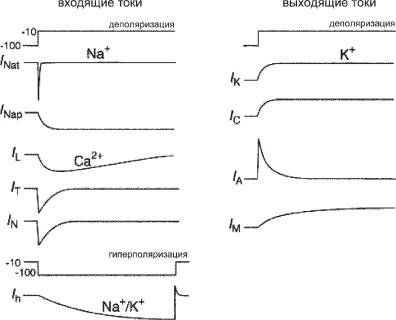
Ðèñ. 6.36. Схематическое отображение потенциал-зависимых катионных токов клеточных мембран мозга млекопитающих. Токи активируются скачком потенциала от -100 до -10 мВ (деполяризация) или от -10 до -100 мВ (гиперполяризация).
INa,t (transient). Активируется и инактивируется в течение нескольких миллисекунд. Все исследованные на сегодня нейроны ЦНС обладают высокоамплитудным INa,t, в то время как INa,p существенно меньше по амплитуде. Быстрая активация и инактивация делает этот ток
идеальным для выполнения роли генерации ПД.
INa,p (persistent, на рис. 6.34 обозначен как INa+,slow). Этот ток быстро активируется при деполяризации, но медленно инактивируется. По-
добно синаптическому потенциалу, поддерживает мембранный потенциал около порога генерации ПД. Дополнительная деполяризация усиливает ответ нейрона на приходящее возбуждение.
Ñà2+-òîêè
В настоящее время выделяют несколько Са2+-токов, различающихся по молекулярной структуре пропускающих их каналов, пороговым
179