

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdf6.6.1. Быстрые ВПСП
Ацетилхолиновые (АХ) ВПСП вызываются активацией íèêî-
тинового АХ-рецептора, который открывает канал для катионов Na+, K+ è Ca2+. Потенциал реверсии этого тока около –5 мВ. Эти ВПСП регистрируются в нейронах ЦНС, спинном мозге (в клетках Реншоу), вегетативной нервной системе (в постганглионарных нейронах), ске-
летных мышцах.
Глютаматные ВПСП вызываются активацией трех типов глютаматных рецепторов, имеющих свои специфические агонисты: (1) AMPA,
(2)каиновая кислота и (3) NMDA.
AMPA- и каиновые рецепторы открывают каналы для катионов
Na+, K+ и иногда Ca2+. Потенциал реверсии этого тока около –0 мВ. При ПП (-75 мВ) NMDA-рецептор блокирован ионами Mg2+; ïðè
деполяризации блок снимается, и через канал течет Na+/K+/Ca2+-ток, вызывающий продолжительную деполяризацию. Активация NMDAрецептора может вызвать продолжительный ток, лежащий в основе долговременной потенциации.
6.6.2. Быстрые ТПСП
ÃÀÌÊ-ТПСП вызываются активацией ÃÀÌÊA-рецепторов, которые открывают канал для Cl-. Потенциал реверсии Cl--тока составляет около –75 мВ. Cl--ток обеспечивает быстрое торможение, например, пирамидных клеток коры и гиппокампа, вызываемое тормозными ин-
тернейронами через коллатеральные тормозные пути.
ÃÀÌÊA- è ÃÀÌÊÑ-рецепторы управляют Cl--проводимостью в дендритах колбочковых и палочковых биполяров сетчатки позвоночных,
соответственно.
Глициновые ТПСП вызываются активацией глициновых рецепторов, которые так же, как ÃÀÌÊÀ(Ñ)-рецепторы, открывают канал для Cl-.
6.6.3. Медленные тормозные эффекты
Увеличение Ê+-проводимости обеспечивается действием многих медиаторов, например, ацетилхолина, норадреналина, серотонина, ГАМК, дофамина, аденозина, соматостатина, энкефалинов. Пирамид-
ные клетки гиппокампа отвечают на серотонин, ГАМК (при участии метаботропных ÃÀÌÊÂ-рецепторов) и аденозин увеличением Ê+-ïðî-
водимости. Функциональное значение увеличения К+-проводимости состоит в торможении, в результате которого вероятность генерации ПД в нейроне уменьшается.
150
Уменьшение Ca2+-проводимости. АХ, норадреналин, серотонин и ГАМК могут уменьшать Ca2+-ток в пресинаптических мембранах. Уменьшение этого тока рассматривается как отрицательная обратная связь, которая регулирует уровень выделившегося в синаптическую щель медиатора. Поскольку выделение медиатора обеспечивается входом Ca2+ в аксонную терминаль, уменьшение входящего Ca2+-тока уменьшает количество выделившегося медиатора.
6.6.4.Медленные возбудительные эффекты (уменьшение Ê+-проводимости)
Âмембранах нейронов обнаружены потенциал-зависимые К+-òîêè
(параграф 6.8.10), которые блокируются рядом медиаторов. Уменьшение IAHP (Ê+-ток следовой гиперполяризации) показано
при действии ряда медиаторов — норадреналина, ацетилхолина, серотонина, гистамина, глютамата. Этот ток регулируется через систему вторичных посредников при участии цАМФ и обеспечивает адаптацию частоты спайковых разрядов, т.е. постепенное ее уменьшение до устранения ПД. При уменьшении IAHP адаптация устраняется (ослабевает), что приводит к продолжительной импульсации нейронов в течение всего времени действия стимула. Функциональное значение этого тока состоит в увеличении отношения «сигнал-шум» в ответе нейрона на
стимул.
Уменьшение IÌ (мускариновый К+-ток) обеспечивается мускариновыми АХ-рецепторами. Этот ток так же, как и IAHP , участвует в адаптации спайковых разрядов, активируется при деполяризации выше
–65 мВ и, в отличие от IAHP , не проявляется при ПП. IÌ уменьшается при активации различных рецепторов, включая серотонинэргические,
глутаматэргические, а также некоторые пептидные рецепторы.
6.7. Проведение локальных потенциалов
Пассивные электрические свойства нейронов (сопротивление и емкость мембраны, а также сопротивление цитоплазмы) играют важную роль в распространении электрических потенциалов по нейронным сетям. В периферических рецепторах потенциалы возникают при действии раздражителей различной физической природы (рецепторные потенциалы), в постсинаптических мембранах потенциалы возникают в результате ионной проводимости при синаптической передаче (постсинаптические потенциалы). Амплитуда таких потенциалов градуально изменяется в зависимости от амплитуды токов, протекающих
151
через мембраны периферических рецепторов и нейронов. В органах чувств через градуальные потенциалы происходит генерация импульса в ответ на сенсорный стимул; в аксоне они способствуют распространению нервного импульса; в постсинаптических нейронах такие потенциалы возникают на многочисленных синаптических входах и суммируются, определяя тем самым вероятность генерации ПД в нейроне.
В данном разделе рассмотрено распространение потенциалов по нервным волокнам с постоянным диаметром, т.е. по цилиндрическим проводникам, и в условиях, когда в мембранах не активируются потен- циал-зависимые каналы, изменяющие сопротивление мембраны.
6.7.1. Кабельные свойства нервных и мышечных волокон
Нервное волокно состоит из тех же компонентов, что и обычный электрический кабель. Роль проводящей среды в нервном волокне выполняет раствор солей, обладающий плохой проводимостью в отличие от металлических проводников. Мембрана клетки является изолятором, но из-за малой толщины обладает высокой емкостью. Из-за таких электрических особенностей потенциал, приложенный к нервному волокну, распространяется только на незначительное расстояние. С некоторыми упрощениями продольное распространение тока по волокну подчиняется закону Ома, согласно которому ток i, проходя через сопротивление r, создает напряжение v = ir.
6.7.1.1. Входное сопротивление и постоянная длины
Прямое электрическое раздражение волокна вызывает потенциал, который пассивно распространяется по волокну, как по кабелю. По мере удаления от места возникновения амплитуда потенциала падает по экспоненциальному закону (рис. 6.14). Эквивалентная электрическая схема мембраны на рисунке 6.15 объясняет падение амплитуды потенциала. Формально мембрана состоит из дискретных контуров cm- rm и дискретных продольных сопротивлений ri. Сопротивление мембраны rm представляет собой сопротивление стенки цилиндра, а продольное сопротивление ri соответствует внутреннему сопротивлению участка аксоплазмы между цилиндрами.
По мере увеличения продольного сопротивления (в результате последовательной суммации ri) ток уменьшается, так как при последовательном соединении сопротивления складываются, и по закону Ома ток будет уменьшаться (I = U/R). Поскольку все сопротивления rm в контуре одинаковы, потенциалы на них будут пропорционально умень-
152
шаться, так как через них проходит уменьшающийся ток. В результате трансмембранная разность потенциалов уменьшается по мере удаления от источника.
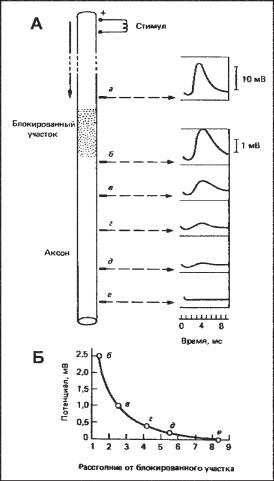
Ðèñ. 6.14. (А) Пассивное распространение возбуждения по аксону кальмара. Участок волокна блокировали локальным охлаждением, чтобы предотвратить генерацию ПД. Потенциал преодолевает блокированный участок, существенно уменьшаясь по амплитуде. При распространении далее по волокну амплитуда потенциала градуально снижается. (Б) График зависимости амплитуды потенциала от расстояния от блокированного участка.
153
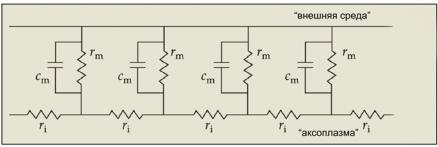
Ðèñ. 6.15. Эквивалентная электрическая схема мембраны. Электрические характеристики локального участка волокна: ri — внутреннее продольное сопротивление, rm — поперечное сопротивление, cm — емкость мембраны.
Падение амплитуды потенциала носит экспоненциальный характер, так что потенциал (Vx) при данном значении расстояния õ равен:
Vx = V0e-x/λ
Максимальный потенциал V0 определяется величиной стимулирующего тока. Уменьшение потенциала определяется константой λ, называемой постоянной длины волокна. λ — это расстояние, на котором потенциал снижается в å раз, т.е. до 37% от максимального значения.
6.7.1.2. Сопротивление мембраны и продольное сопротивление
Постоянная длины волокна λ зависит как от rm , òàê è îò ri :
λ = (rm/ri)½
Исходя из формулы, λ возрастает с увеличением сопротивления мембраны, препятствующего утечке тока во внеклеточное пространство, и, напротив, снижается с увеличением внутреннего сопротивления, которое затрудняет продольное протекание тока по аксоплазме.
Еще одна важная кабельная характеристика проводника — входное сопротивление мембраны — также зависит от обоих параметров:
rinput = 1/2(rm/ri)1/2
Коэффициент 1/2 в формуле объясняется тем, что аксон простирается в двух направлениях от места инъекции тока и каждая его половина обладает своими сопротивлениями. Измерив в эксперименте rinput è λ, можно рассчитать rm è ri.
154
6.7.1.3. Удельное сопротивление
Рассчитанные значения rm è ri, характеризуют сопротивление модельного цилиндрического сегмента аксона длиной 1 см. Однако эти характеристики не предоставляют точной информации о сопротивлении мембраны и аксоплазмы, поскольку последние зависят от размера волокна. Сопротивление мембраны нервного волокна обратно зависит от общего числа ионных каналов, которое определяется их плотностью и площадью поверхности сегмента аксона.
Чтобы сопоставить между собой мембраны разных волокон, нужно знать величину удельного сопротивления Rm, которое отражает сопротивление мембраны площадью 1 см2 и измеряется в Ом см2. Отрезок аксона длиной 1 см и радиусом α обладает площадью поверхности 2πα ñì2. Сопротивление мембраны rm такого отрезка составляет Rm/2πα, откуда Rm=2πα rm.
В большинстве типов нейронов Rm определяется главным образом К+- è Cl–-проводимостями мембраны в покое. Среднее значение Rm, аксона омара составляет около 2000 Ом см2. Значения Rm, полученные на других объектах, варьировали в диапазоне от 1000 Ом см2 äî 50000 Îì ñì2 в зависимости от плотности ионных каналов.
Удельным сопротивлением аксоплазмы Ri считают внутреннее продольное сопротивление сегмента аксона длиной 1 см и площадью сече- ния 1 см2.
Поскольку с увеличением площади сечения цилиндра его продольное сопротивление уменьшается, то ri = Ri/πα2, откуда Ri = πα2ri.
Величина Ri измеряется в Ом см, и ее значение для аксона кальмара составляет около 30 Ом см. Ri обратно зависит от концентрации ионов в аксоплазме. У животных с более низкой ионной концентрацией в аксонах Ri составляет около 125 Ом см (у млекопитающих) и около 250 Ом см (у земноводных).
6.7.1.4. Влияние диаметра кабеля на его характеристики
На основе уравнений, приведенных выше, можно определить входное сопротивление волокна как:
rinput = 1/2( RmRi /2π2α3)1/2
Следовательно, входное сопротивление волокна снижается как 3/2 степень его радиуса.
155
Так же постоянная длины волокна λ определяется соотношением его поперечных и продольных удельных сопротивлений (Rm è Ri) и его радиусом α:
λ = (αRm/2Ri)1/2
Таким образом, λ возрастает с увеличением радиуса. Измеряя сопротивления и радиусы разных волокон, можно сопоставить их кабельные свойства. Так, значения постоянной длины волокна аксона кальмара (диаметром до 1 мм), мышечного волокна лягушки (диаметром 50 мкм) и нервного волокна млекопитающих (диаметром 1 мкм) составляли, соответственно, 13, 1,4 и 0,3 мм.
Итак, кабельные свойства проводника rinput и λ определяют амплитуду сигнала, генерируемого нервным волокном, и его декремент с расстоянием. Например, при равных условиях синаптического воздействия величи-
на ВПСП в тонком дендрите (с более высоким rinput) будет больше, чем в толстом. С другой стороны, в толстом дендрите из-за большего значения λ
потенциал будет меньше затухать с расстоянием, чем в тонком.
6.7.1.5. Постоянная времени
Мембрана обладает свойством емкости, накапливая заряд на своей внешней и внутренней поверхности. Как у конденсатора, два слоя жидкости по обе стороны мембраны являются обкладками, а сама мембрана представляет собой изолирующую прослойку. При толщине мембраны 5 нм она способна накапливать достаточно большой заряд. Обычно емкость мембраны нервных клеток составляет 1 мкФ/см2. Емкость мембраны приводит к тому, что фаза роста амплитуды потенциала, возникающего на мембране в результате приложения к ней тока, развивается по экспоненте в соответствии с уравнением:
Vt = V0(1-e-t/τ),
ãäå t — время от начала импульса, Vt — амплитуда потенциала в момент времени t, V0 — максимальная амплитуда потенциала, τ — постоянная времени, равная произведению удельного сопротивления и удельной емкости мембраны (RmCm). Постоянная времени отражает время, за которое потенциал достигает 63% (1–1/е) от максимального значения амплитуды. Спад потенциала после выключения также происходит по экспоненте с той же постоянной времени.
Постоянная времени мембраны не зависит от размера клетки или волокна. Увеличение радиуса (и площади поверхности мембраны) приводит не только к увеличению удельной емкости, но и к
156
снижению удельного сопротивления мембраны. При такой обратной зависимости этих параметров от размера волокна τ, как их произведение, (RmCm) не меняется. Постоянная времени, наряду с входным сопротивлением и постоянной длины, является параметрам, от которого зависят свойства аксона как проводника. Диапазон значений постоянной времени в различных типах нервных и мышечных клеток составляет от 1 до 20 мс.
Емкость мембраны приводит к замедлению поляризации мембраны в результате действия на нее тока. Поскольку при параллельном соединении мембранных модулей их суммарная емкость увеличивается, то по мере удаления от места раздражения время заряда емкости также увеличивается. Поэтому чем дальше от места раздражения развивается потенциал, тем τ больше, и фазы роста и спада потенциалов замедляются.
Еще одно существенное влияние мембранной емкости на потенциалы заключается в том, что кратковременные токи, активирующие мембрану, распространяются на более короткие расстояния, чем длительные. В случае продолжительного сигнала потенциал достигает своего максимального значения и емкость заряжается полностью. В результате пространственное распределение потенциала определяется только сопротивлениями мембраны и цитоплазмы (Vx = V0e-x/λ). При действии коротких сигналов (например, синаптический потенциал) ток прекращается еще до того, как емкость успевает полностью зарядиться. Это выражается в уменьшении расстояния, на которое потенциал распространяется вдоль волокна.
6.7.2. Влияние кабельных свойств на возбудимость нейронов
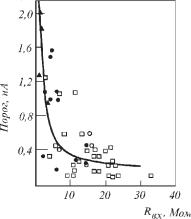
Кабельные свойства нейронов и их отростков во многом предопределяют их реакции на синаптические воздействия. Величина порогового тока у разных нейронов варьирует в широком диапазоне — 10-7–10-10 А. Оказалось, что величина порогового тока обратно зависит от входного сопротивления мембраны нейрона. Зависимость порогового тока от сопротивления описывается гиперболой (рис. 6.16), из чего следует, что пороговый ток определяется входным сопротивлением, а пороговый потенциал является постоянным для электровозбудимых мембраны нейронов. Таким образом, эффективность синапсов, вызывающих токи в мембранах, зависит от входного сопротивления дендрита.
Входное сопротивление волокна увеличивается с уменьшением его радиуса, что было подтверждено специальными измерениями (рис. 6.17).
157
Из этого следует, что более тонкие дистальные дендриты обладают большим входным сопротивлением и, соответственно, меньшие токи могут сдвигать мембранный потенциал к пороговому уровню генерации ПД. Поэтому эффективность синапсов, активирующих дистальные дендриты, может значительно превышать эффективность синапсов на близких к соме дендритах. Из этого также следует важный вывод о том, что синаптические влияния на мелкие нейроны больше, чем на крупные.
Ðèñ. 6.16. График зависимости порога возбуждения мембран нейронов моторной коры кошки от входного сопротивления. Экспериментальные точки аппроксимированы методом наименьших квадратов.
6.7.3. Проведение потенциалов по сложным нейронным отросткам
Самой простой моделью нейрона является гипотетический сфери- ческий нейрон, который представлен множеством параллельно соединенных RC-элементов (рис. 6.18). Все участки в такой модели включе- ны параллельно, поскольку сопротивление цитоплазмы и внешней среды пренебрежимо мало по сравнению с сопротивлением мембраны. Принимая во внимание такое упрощение, эквивалентная электрическая схема сферического нейрона совпадает со схемой единичного элемента мембраны. При стимуляции такого простого RC-элемента по-
тенциал на мембране будет изменяться по экспоненте с постоянной времени τ = RC.
158

Ðèñ. 6.17. Входные сопротивления в теле и дендритах мотонейрона кошки. Числа указывают входное сопротивление (мОм) при удельном сопротивлении мембраны 50 Ом см.
Ðèñ. 6.18. Эквивалентная электрическая схема элемента мембраны (А) и сферического нейрона. Ñ — емкость мембраны, R — сопротивление мембраны, R’ — сопротивление протоплазмы R’’ — сопротивление наружной среды, Ì — мембрана. У шаровидной клетки R’ è R’’ << R.
159