

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfэффекты, как привыкание, фасилитация или аналог условного ответа, требуют участия кальция: в бескальциевой среде предъявление даже большого количества стимулов не вызывало каких-либо следовых преобразований реакций (рис. 13.17).
Кальциевые осцилляции — это периодические выбросы внутриклеточного кальция, а кальций-зависимые внутриклеточные процессы регулируются частотой кальциевых осцилляций (Tomida et al., 2003). Хотя функциональное значение этих кальциевых осцилляций общепризнано, механизмы, лежащие в основе их запуска и развития, требуют дальнейшего изучения. Предполагается, что роль пейсмекера в генерации кальциевых осцилляций могут выполнять передвижения ионов кальция между эндоплазматическим ретикулкулумом и митохондриями (Ishii et al., 2006). Интересно, что кальциевая природа пейсмекерных осцилляций, по-видимому, является «наследием прошлого»: в работах на простейших показано, что при дефиците кальция поведение одноклеточных резко изменяется, они теряют подвижность (Wassenberg, Clark, 1997), а осцилляторы связаны с кальциевой проводимостью мембраны (Eckert. and Naitoh, 1970).
Нейрофизиологические исследования показывают, что пейсмекерные потенциалы имеют следующие свойства: 1) пейсмекер обладает высокой чувствительностью к изменениям уровня мембранного потенциала; 2) актуализация пейсмекера связана с достижением определенного уровня мембранного потенциала; 3) пейсмекер взаимодействует с синаптическими потенциалами и может быть включенным (или выключенным) через определенный синаптический вход; 4) пейсмекерный механизм может запускаться действием биологически активных веществ в сверхмалых концентрациях; 5) пейсмекер участвует в обуче- нии нейронов.
13.4. Пейсмекер и функции
Функциональное назначение эндогенных осцилляций различно, поэтому и рабочие режимы нейронов, требующие участия пейсмекерной активности, также отличаются. Для одних клеток активное проявление пейсмекерного механизма является нормой, например, для командного нейрона эндогенная ритмика — это способ реализации внутренней программы, определяющей будущую двигательную активность, для нейрона-модулятора — инструмент регуляции состояния целых нейронных популяций. Включаясь в определенных усло-
360
виях, такие пейсмекерные нейроны приводят ансамбли клеток к гармоничному взаимодействию и, следовательно, к адаптивному поведению живого существа. Другие же нейроны реализуют свой эндогенный осциллятор только в момент гибели, подавая сигнал об этом в межклеточную среду.
Результаты опытов на одноклеточных и многоклеточных показывают, что усложнение структуры и организации нервной системы, связанное с появлением синаптической передачи, не привело к вырождению или обеднению функций пейсмекерной активности, скорее, наоборот — к усилению ее значения. Эндогенная ритмика, сформировавшаяся под влиянием циклических процессов окружающей среды еще на ранних этапах появления живых организмов, наиболее надежно и адекватно «вписывает» сложные системы в быстро меняющуюся обстановку. Не случайно ход биологических часов животных различного эволюционного уровня реализуется пейсмекерными нейронами (у животных, имеющих развитый головной мозг, нейронами супрахиазменного ядра) (Hahn et al., 2006, Leznik and Llinás, 2005, Meijer J. H., Rietveld, 1989, Mieda, 2006, Rabbah P, Nadim, 2007). Поломки в функциях пейсмекеров различных мозговых структур могут приводить к заболеваниям, например, нарушение взаимодействия пейсмекеров внешнего сегмента бледного ядра с пейсмекерами базальных ядер приводит к развитию тремора (Rabbah, Nadim, 2007), процедурное обучение ухудшается при полушарных операциях на мозжечке (Gómez-Beldarrain, 1998), изменение условий для взаимодействия пейсмекеров дорзомедиальных гипоталамических ядер приводит к реорганизации пищевого поведения (Mieda et al., 2006). Дофаминовые нейроны среднего мозга, активность которых существенна для выполнения когнитивных и двигательных задач, генерируют пейсмекерные потенциалы, включение-выключение которых зависит от активности синаптических входов. Нарушение функции этих клеток приводит к развитию неврологических и психических заболеваний, таких, как болезнь Паркинсона, шизофрения и наркозависимость (Johnson et al., 2005). Нарушение функции пейсмекеров продолговатого мозга, участвующих в регуляции дыхания, вызывает развитие удушья (Ramirez JM, Garcia, 2007). В научной литературе настоящего времени накоплен колоссальный материал, демонстрирующий многогранность проявления пейсмекера в самых разных функциях живого существа и на различных этапах его жизни — от зарождения до смерти.
361
Пейсмекер — начало жизни
Внутренние кальциевые волны и осцилляции рассматриваются как универсальное свойство активности клетки. Как важнейший вторич- ный посредник, кальций запускает целый ряд клеточных реакций, вклю- чая сокращения, секрецию и экспрессию генов. Некоторые факторы чрезвычайно важны для определения того, будет ли это единичный всплеск изменения концентрации кальция или же будут инициироваться волны. Эксперименты показали, что при оплодотворении в яйцеклетке животных и растений любого вида возникает единичное или транзиторное изменение содержания кальция (рис. 13.20). У эукариотических клеток кальциевая волна стартует с повышения исходного содержания кальция в ограниченной области клетки (пейсмекерном локусе), откуда затем распространяется, приводя к глобальной кальциевой волне. У яйца глобальная кальциевая волна всегда инициируется на поверхности и затем распространяется по поверхности или через всю цитоплазму. После инициации кальциевой волны ее распространение связано с активацией кальцием же Са2+-каналов на фронте волны. На скорость распространения кальциевой волны может влиять пространственная организация мест реализации Са2+ и скорость пассивной диффузии Са2+.
У яйца Са2+ волны запускаются вторжением спермы, и в основном она является результатом реализации Са2+ из внутриклеточных хранилищ через посредство инозитол-1,4,5-трифосфата. Механизм, лежащий в основе продукции этого вещества в клетке, остается туманным и в настоящее время активно обсуждается. Аналогично природа фактора, который содержится в сперме и под влиянием которого начинается высвобождение Са2+, остается непонятной. Некоторые исследователи предполагают, что это может быть протеин, возможно, форма фосфолипазы С или ее активатор. Кажется вполне вероятным предположение о том, что у большинства видов вхождение спермы вносит этот фактор в яйцо и этот фактор прямо или косвенно вызывает синтез (или высвобождение) инозитол-1,4,5-трифосфата (Dumollard et al., 2002).
Первая волна, которая рассматривается как «Са2+ волна оплодотворения», обычно самая большая и длительная, и у некоторых видов она сопровождается повторными волнами кальция снижающейся амплитуды и меньшей длительности. Пейсмекеры Са2+ волны возбуждают колебания в течение 20-30 мин. у асцидий, 45-60 мин. у некоторых моллюсков, несколько часов у млекопитающих, и они перестают действовать только в конце клеточного мейотического цикла (кроме мле-
362
копитающих, у которых они прекращаются через несколько часов после завершения мейоза во время формирования пронуклеуса). Пейсмекерный локус может фиксироваться на поверхности или же передвигаться, когда поверхность реорганизуется для подготовки к развитию (рис. 13.20). Са2+ волны пейсмекера локализуются или в области повышенной чувствительности к инозитолу, или же располагается вблизи локального источника инозитола. Каким образом субклеточная организация высвобождения Са2+ может создавать стабильные волны пейсмекера в яйце? Это вопрос, на который еще нет ответа.
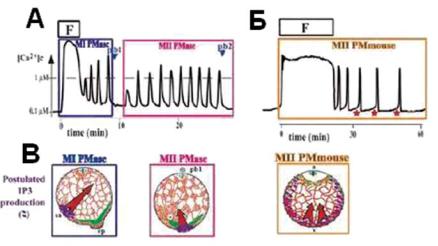
Ðèñ. 13.20. Пейсмекерные волны Ca2+ на кортикальных слоях яйцеклетки асцидии и мыши. А — инициированные сперматозоидом волны Са2+ у асцидии: мейотические волны кальция, состоящие из волны, возникшей при оплодотворении (F), сопровождаемые повторяющимися Са2+ волнами, инициированными двумя пейсмекерами (MI PMasc и MII Pmasc). Б — яйцеклетка мыши оплодотвряется на метафазе II и поэтому вовлекает только пейсмекер МII (MII PMmouse). После волны, возникающей при оплодотворении (F), на- чинающейся от пункта вторжения спермы, повторные волны исходят из вегетального кортекса яйца. Каждой кальциевой волне предшествует пейсмекерный кальциевый подъем (красные звездочки). В — схематичное изображение организации сети эндоплазматического ретикулума (ER) (красное) и митохондрий (зеленое) яйцеклетки асцидии (MI Pasc и MII Pasc) и мыши (MII Pmouse). Красные стрелки показывают направление движения волн, когда постулируется определенное место продукции Ins(1,4,5)P3 (фиолетовые точки). (по R. Dumollard, J. Carroll, G. Dupont and Christian Sardet. Calcium wave pacemakers in eggs Journal of Cell Science 115, 3557-3564 2002).
363
У яиц млекопитающих и у асцидий большие, запускаемые спермой волны кальциевой активации пересекают яйцо при оплодотворении, сопровождаемые серией периодического повышения внутриклеточной кальциевой концентрации. Показано, что у яиц асцидии эти периодические кальциевые увеличения появляются в форме волны, источник которой перемещается примерно в течение 5 мин. в пейсмекерную область. На полюсе, богатом уплотнениями эндоплазмати- ческого ретикулума, появляется место локализации пейсмекера, из которого исходят кальциевые волны. Как только источник генерации приобретает стабильную локализацию, кальциевые волны начинают распространяться по поверхности яйца (McDougall A, Sardet C, 1995). У яиц асцидии постактивационные кальциевые волны необходимы для завершения мейоза. Их инициация прекращаются перед его завершением.
Осцилляции концентрации внутриклеточного кальция время от времени запускаются гормоном созревания (как, например, у моллюсков) и обычно наблюдаются после оплодотворения, которое происходит на разных стадиях мейотического цикла клетки. У большинства изученных таким образом яиц кальциевые осцилляции продолжаются до конца мейоза и перед формированием мужского и женского пронуклеуса. Эта способность зависит от чувствительности InsP3 каналов и от проницаемости плазматической мембраны для экстраклеточного кальция. У яйца, которое испытывает цитоплазматичесеую реорганизацию при оплодотворении, повторные кальциевые сигналы являются волнами, исходящими от локализациии кортикального локуса, который становится пейсмекером кальциевых волн. У асцидий было идентифицировано место инициации повторных калциевых волн как скопление прослоек эндоплазматического ритикулума между плазматической мембраной и скоплением митохондрий (Sardet et al., 1998).
Пейсмекер — часовая стрелка эндогенных ритмов
Циркадианные часы — широко распространенный клеточный механизм, который лежит в основе различных ритмических функций организмов от бактерий до грибов, от растений до животных, и организует их метаболизм, чтобы приспособиться к смене дни и ночи на Земле ( Pittendrigh, 1993, Young MW, Kay SA, 2001). Этот механизм развился у множества живых организмов (McClung, 2006). Правиль-
364
ное функционирование эндогенных циркадных часов дает живому существу возможность предвидеть ежедневные изменения окружающей среды и соответственно вовремя изменять поведенческие и физиологические функции. Все организмы сохраняют постоянными большое количество физиологических переменных (сон-бодрствование, локомоторную активность, температурную регуляцию, поступление еды и воды, уровень циркулирующих гормонов) под контролем циркадных часов. Существуют хорошо известные последствия прерывания циркадианной функции. Метаболизм, репродукция и даже долгожительство могут затрагиваться в неблагоприятную сторону, если определение времени дня изменяется на молекулярном уровне. Пожалуй, меньше можно сказать относительно мозга и идеи о том, что измененная функция часов может участвовать в поведенческих, нейрологических и психических растройствах. Циркадные ритмы проявляют все виды живых существ. Ритмичность поведения сохраняется даже в постоянных условиях, при периодах немного короче или длиннее суток, тем самым показывая, что осцилляции представляют эндогенную временную программу. У животных дневная активность измеряется обычно как активность, противопоставляемая отдыху, но существует громадное количество других параметров от поведенческого уровня до экспрессии генов, которые осциллируют с циркадным периодом. Растения проявляют множество феноменов циркадных ритмов, включая движения листьев, скорость роста, экспрессию механизмов фотосинтеза. Грибы Neurospora crassa создают споры каждые 22 часа в полной темноте, а прокариотические организмы — цианобактерии Synechococcus осуществляют несовместимые процессы фотосинтеза и азотной фиксации в разное время дня, решая проблему использования времени, а не пространства, как это деляют другие цианобактерии. Устойчивые ритмы наблюдали у одноклеточных (LakinThomas and Brody, 2004), у диссоциированных нейронов, выделенных из супрахиазменного ядра (Welsh et al, 1995), даже фибробласты крысы в культуре ткани демонстрировали циркадный ритм экспрессии гена (Balsalobre et al, 1998). Cуточные биологические ритмы широко распространены в природе и связаны с такими сигналами окружающей среды, как свет и температура, синхронизируя внутреннее время с 24-часовым вращением Земли.
Разрушение генов циркадных ритмов у мышей-мутантов влияет на их поведение. Наконец, предполагая вклад циркадианной регуляции в
365
симптоматологию, исследователи изучали связь между полиморфизмом часовых генов, поведенческими особенностями и расстройствами у человека. Постепенно формируется представление о том, что генная разрегуляция часового механизма влияет на широкий спектр болезненных расстройств ЦНС.
При постоянном уровне затемнения и температуре организм может проявлять один или несколько циркадных ритмов в осуществлении таких функций, как локомоция, чувствительность к ядам или специфическая активность индивидуальных энзимов. Период таких ритмов обычно равен примерно 24 часам. Как правило, он точный и температурно-компенсируемый. В качестве циркадных систем могут рассматриваться как целые организмы, так и отдельные клетки, чьи выходные проявления циркадных ритмов (непосредственно наблюдаемые) должны быть результатом периодичности в работе физиологической системы. Ясно, что общая система (целая) включает в себя, кроме того, управляющий элемент. Сам он может быть или не быть периодическим. Не обязательно, чтобы его осцилляции в точности соответствовали периодичности всей
системы (Pittendrigh, 1967).
Циркадианные ритмы простейших. Долгое время считали, что у прокариотов невозможно экспрессирование циркадных ритмов. Однако недавно такие биологические часы были обнаружены у некоторых видов цианобактерий. Цианобактерии являются простейшими из известных организмов, которые проявляют циркадные ритмы (Iwasaki and Kondo, 2004). Эти эндогенные хранители времени у цианобактерий контролируют экспрессию генов. Даже у цианобактерий, которые выращиваются в культуре с удвоенным течением времени (более 1 за 24 часа) циркадные часы контролируют экспрессию генов и деление клеток. В работе Johnson CH, Golden SS, Ishiura M, Kondo T. (1996) изолировали мутации циркадианного пейсмекера у цианобактерий и определяли локусы, ответственные за изменение периодичности фенотипа.
Выяснилось, что суточный ритм цианобактерий определяется циклическими реакциями присоединения фосфатных групп к определенным участкам группы белков. Исследователям удалось установить механизм работы простейшей бактериальной системы из трех белков: KaiA, KaiB и KaiC. Их взаимодействие приводит к циклическим изменениям каждого из них. Самое интересное зак-
366
лючается в том, что полный цикл превращений занимает одни сутки. При этом обнаруженные изменения не зависят от внешних воздействий, что в полной мере объясняет возникновение циркадианных ритмов. KaiA, KaiB, и KaiC — основные элементы циркадных часов у цианобактерий Synechococcus elongatus PCC 7942. KaiC репрессирует свою собственную (kaiBC) экспрессию, а KaiA усиливает экспрессию kaiBC. Поэтому KaiC и KaiA были названы, соответственно, негативным и позитивным факторами, функционирующими в обратной циркадной связи, регулирующей экспрессию kaiBC. О регуляции часовых генов через обратную связь сообщалось применительно к часовой системе эукариотов, таких, как дрозофилы, нейроспора, Arabidopsis и позвоночные (Young and Kay, 2001), хотя никаких дальнейших сходств между Kai протеинами и протеинами часов эукариотов не наблюдалось. Интенсивный генетический анализ, предпринятый в течение последних лет, открыл многие компоненты и молекулярные механизмы, позволяющие сравнивать эти часы. Хотя авторегуляция генетической системы является основным свойством в устройстве всех часов, свод данных поддерживает их независимое эволюционное происхождение в разных царствах (Young, Kay, 2001).
Один из самых важных вопросов исследования биологических ритмов заключается в следующем: как стабильные и точные 24-часовые осцилляции генерируются на молекулярном уровне. Хотя казалось, что возрастающая сложность является ключом, но недавние сообщения заставляют предполагать, что около 24-часовые ритмы генерируются всего четырьмя протеинами, инкубируемыми в пробирке (Roenneberg et al., 2008).
KaiA, KaiB, и KaiC — это основные протеины циркадных часов у цианобактерий. Считается, что цикл фосфорилирования KaiC, который происходит in vitro после смешения трех протеинов и АТФ — это и есть основной осциллятор , управляющий циркадной системой. Был проанализирован временной профиль комплексов, который формируется между тремя комплексами протеинов (рис. 13.21). На фазе фосфорилирования активируется протеин KaiA и повторно связывается с KaiC, чтобы запустить фосфорилирование KaiC. В работе исследована последовательность событий, связанных с экспрессированием белков, которые участвуют в циркадном ритме (Kageyama et al., 2006).
367
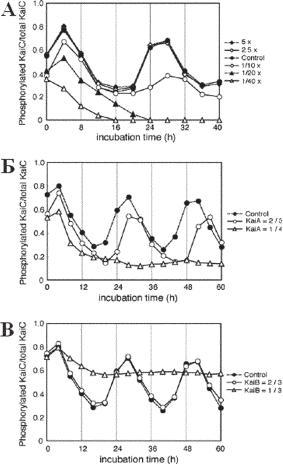
Ðèñ. 13.21. Ритм фосфорилирования KaiC при различных концентрациях протеинов Kai. В.Относительная интенсивность сигналов фосфорилируемого KaiC, определяемая посредством денситометрии, показывает время инкубации in vitro. С-Ритм фосфорилирования KaiC в условиях снижения концентрации KaiA (2/3 или 1/4 концентрации стандартного раствора. Обозначения прежние. D— Ритм фосфорилирования KaiC при снижении концентрации KaiB (2/3 или 1/3 концентрации стандартной смеси. (по Molecular Cell 23, 161–171, July 21, 2006 Сyanobacterial Circadian Pacemaker: Kai Protein Complex Dynamics in the KaiC Phosphorylation Cycle In Vitro Hakuto Kageyama, 1 Taeko Nishiwaki, 1 Masato Nakajima,1 Hideo Iwasaki, 1, 2 Tokitaka Oyama, 1 and Takao Kondo1).
368
Двумя чрезвычайно важными биологическими событиями являются циркадианные ритмы и цикл деления клетки. Деление клетки происходит у всех растущих организмов, а циркадианные ритмы обнаружены у всех организмов от прокариотических цианобактерий до эукариотов, включая простейших, грибы, растения, животных и человека. Фундаментальными характеристиками циркадианных ритмов, которые определяют и отличают их от других периодических феноменов живых организмов, являются: 1) они эндогенны и генетически определены; 2) ритмы продолжаются около 24 часов при постоянных условиях, которые опосредованы условиями окружающей среды; 3) продолжительность периодов компенсируется изменениями температуры окружающей среды в широком ряду физиологически релевантных температур (Mori and Johnson, 2001).
У цианобактерий деление клетки регулируется циркадными ча- сами. Удаление гена циркадных часов, KaiC, уничтожает ритмы экспрессии гена и расписание деления клетки. Сверхэкспрессия гена fisZ поддерживает деление клетки, но не рост, что побуждает клетку увеличиваться как филамент, без деления. Неделимая клетка все еще способна проявлять грубые приблизительные экспрессии гена циркадных ритмов. Эти результаты показывают, что циркадианное расписание системы является независимым от ритмического деления клетки и вместе с другими результатами заставляет предполагать, что циркадианная система цианобактерий стабильна и хорошо сохраняется при влиянии целого ряда внутриклеточных условий (Mori et al., 2001).
Таким образом, было показано, что экспрессия часовых генов и деление клетки осуществляется разными механизмами. Но биологи- ческие часы организуют общую регуляцию экспрессии генов и контролируют расписание деления клетки. Были идентифицированы протеины, которые могут вовлекаться в начальные пути этих процессов. Мутационный скрининг позволил определить три часовых гена, которые организованы в генный кластер. Структура цианобактериальных часовых протеинов, их фосфорилирование и регуляция также описаны. Новая модель сердцевины часового механизма цианобактерий заставляет предполагать, что ритмические изменения состояния хромосомы лежат в основе ритмов экспрессии генов. Mixed-strain эксперименты показали, что этот хранитель времени обеспечивает адаптивное значе- ние, когда разные «племена» конкурируют друг с другом (Johnson, 2004).
369