

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdf170.Ramirez JM, Garcia Iii A. Medullary Pacemaker Neurons are Essential for Both Eupnea and Gasping in Mammals- // The Last Word J Appl Physiol. 2007, Apr. 26, 26, 29.
171.Roenneberg Till 1 and Merrow Martha 2 Circadian Clocks: Translation Lost Current Biology. Vol 15, No 12, PP., 8.
172.Rose T, Gras H, Hörner M. Activity-dependent suppression of spontaneous spike generation in the Retzius neurons of the leech Hirudo medicinalis L. Invert Neurosci. 2006, Dec; 6 (4): 169-76. Epub. 2006, Oct. 31, 14.
173.Ruta V, Jiang Y, Lee A, Chen J, MacKinnon R.. Functional analysis of an archaebacterial voltage-dependent K+ channel. Nature. 2003, Mar. 13; 422 (6928): 180.
174.Sardet C, Roegiers F, Dumollard R, Rouviere C, McDougall A. Calcium waves and oscillations in eggs.. Biophys Chem. 1998, May. 5; 72 (1-2): 131-40., 20.
175.Scott IM, Boulant JA.Brain Res. 1984, Jul. 23; 306 (1-2): 157-63. Dopamine effects on thermosensitive neurons in hypothalamic tissue slices. 2007; 72: 47-55., 9.
176.Shibata S, Moore RY Electrical and metabolic activity of suprachiasmatic nucleus neurons in hamster hypothalamic slices. Brain Res. 1988, Jan. 12; 438 (1-2): 374-8.
177.Simon S. M. and Llinas R.R. Compartmentalization of the submembrane calcium activity during calcium influx and its significance in transmitter release // Biophysical journal, 1985. V. 48 p. 485-497.
178.Soto-Treviño C, Rabbah P, Marder E, Nadim F Computational model of electrically coupled, intrinsically distinct pacemaker neurons J Neurophysiol. 2005, Jul.; 94 (1): 590-604. Epub. 2005, Feb. 23., 34, 40.
179.Sotty F., Danik M., Manseau F., Laplante F., Quirion R.and Williams S.Distinct electrophysiological properties of glutamatergic, cholinergic and GABAergic rat septohippocampal neurons: novel implications for hippocampal rhythmicity J. Physiol (2003), 551.3, pp. 927–943, 14.
180.Sugino K, Tominaga T., Allen R.D. and Naitoh Y. Electrical properties and fusion dynamics of in vitro membrane vesicles derived from separate parts of the contractile vacuole complex of Paramecium multimicronucleatum // The Journal of Experimental Biology. 208, 3957-3969, 34, 40.
181.Sujino M, Masumoto KH, Yamaguchi S, van der Horst GT, Okamura H, Inouye ST. Suprachiasmatic nucleus grafts restore circadian behavioral rhythms of genetically arrhythmic mice. // Curr Biol. 2003, Apr. 15; 13 (8): 664-8, 26.
182.Taddei-Ferretti C, Cordella L.Modulation of Hydra attenuata rhythmic activity: phase response curve // J Exp Biol. 1976, Dec.; 65 (3): 737-51., 26.
183.Thompson RF Are memory traces localized or distributed? Neuropsychologia. 1991; 29 (6): 571-82, 26.
184.Toller GL, Nagy E, Horvath RA, Klausz B, Rekasi Z Circadian expression of Bmal1 and serotonin-N-acetyltransferase mRNAs in chicken retina cells and pinealocytes in vivo and in vitro. J Mol Neurosci. 2006; 28 (2): 143-50., 21.
430
185.Tomida T, Hirose K, Takizawa A, Shibasaki F, Iino M (2003) NFAT functions as a working memory of Ca2+ signals in decoding Ca2+ oscillation // EMBO. J. 22: 3825–3832, 26, 29.
186.Urbano FJ, Simpson JI, Llinás RR. Somatomotor and oculomotor inferior olivary neurons have distinct electrophysiological phenotypes. Proc Natl Acad Sci USA. 2006, Oct 31; 103 (44): 16550-5. Epub. 2006, Oct. 18, 22.
187.Van Houten J. Chemosensory transduction in Paramecium // Europ. J. Protostiol. 1998. V. 34. P. 301-307, 2, 35, 40.
188.Vansteensel MJ, Michel S, Meijer JH. Organization of cell and tissue circadian.
189.Varga V, Hangya B, Kránitz K, Ludányi A, Zemankovics R, Katona I, Shigemoto R, Freund TF, Borhegyi Z The presence of pacemaker HCN channels identifies theta rhythmic GABAergic neurons in the medial septum. J. Physiol. 2008, Aug 15; 586 (16): 3893-915. Epub. 2008, Jun. 19, 16.
190.Victor A. Maltsev, PhD and Edward G. Lakatta, MD Normal heart rhythm is initiated and regulated by an intracellular Calcium clock within pacemaker cells Heart Lung Circ. 2007. October; 16 (5): 335–348., 22.
191.Wassenberg JJ, Clark KD, Nelson DL. Effect of SERCA pump inhibitors on chemoresponses in Paramecium // J. Eukaryot Microbiol. 1997, Nov.-Dec.; 44 (6): 574-81, 27.
192.Wassenberg JJ, Clark KD, Nelson DL. Effect of SERCA pump inhibitors on chemoresponses in Paramecium // J. Eukaryot Microbiol. 1997, Nov.-Dec.; 44 (6): 574-81. 47, 28.
193.Watanabe A, Shibata S, Watanabe S. Circadian rhythm of spontaneous neuronal activity in the suprachiasmatic nucleus of old hamster in vitro. Brain Res. 1995, Oct. 16; 695 (2): 237-9, 10.
194.Welsh DK, Logothetis DE, Meister M, Reppert SM (1995) Individual neurons dissociated from rat suprachiasmatic nucleus express independently phased circadian firing rhythms. Neuron. 14: 697–706, 9.
195.Wittner L, Miles R Factors defining a pacemaker region for synchrony in the hippocampus. J. Physiol. 2007, Nov. 1; 584 (Pt 3): 867-83. Epub. 2007, Sep. 6, 16.
196.Wong R K and M Stewart Different firing patterns generated in dendrites and somata of CA1 pyramidal neurones in guinea-pig hippocampus. J. Physiol. 1992; 457; 675-687.
197.Woolum J. C. and Strumwasser F. The differential effects of ionizing radiation on the circadian oscillator and other functions in the eye of Aplysia (x-rays/molluskan eye) Proc Natl Acad Sci U S A. 1980, Sep.; 77 (9): 5542-6.
198.Wright CL, Burgoon PW, Bishop GA, Boulant JA.Am J Physiol Regul Integr Comp Physiol. 2008, May; 294 (5): R1704-15. Epub. 2008, Mar. 5. Cyclic GMP alters the firing rate and thermosensitivity of hypothalamic neurons., 9.
199.Yamaguchi S, Isejima H, Matsuo T, Okura R, Yagita K. Synchronization of cellular clocks in the suprachiasmatic nucleus. Science. 2003, 302: 1408–1412, 12.
431
200.Yoshimura Hiroshi The Potential of Caffeine for Functional Modification from Cortical Synapses to Neuron Networks in the Brain Current Neuropharmacology, 2005, 3, 309-316, 17.
201.Young MW, Kay SA. Time zones: a comparative genetics of circadian clocks Nat Rev Genet. 2001, Sep.; 2 (9): 702-15, 29.
202.Young, M. W., and Kay, S.A. (2001). Time zones: a comparative genetics of circadian clocks. Nat. Rev. Genet. 2, 702–715., 9.
203.Zhao Y, Boulant JA. Temperature effects on neuronal membrane potentials and inward currents in rat hypothalamic tissue slices. J Physiol. 2005, Apr. 1; 564 (Pt 1): 245-57., 24.
204.Zhi-Gen Jiang*, YuQin Yang*, Zhao-Ping Liut and Charles N. Al Membrane properties and synaptic inputs of suprachiasmatic nucleus neurons in rat brain slices Journal of Physiology (1997), 499.1, pp. 141-159, 9.
205.Zimmerman N. and Menakert M. The pineal gland: A pacemaker within the circadian system of the house sparrow Proc. Natl. Acad. Sci. USA. Vol. 76, No. 2, pp. 999-1003, February 1979.
432
Глава 14. ГЛИЯ: МОРФОЛОГИЯ, ФИЗИОЛОГИЯ И ФУНКЦИИ
Клетки глии распределены по всей нервной системе вокруг нервных клеток и превышают их по численности в 9–10 раз. С момента открытия глии в конце XIX в. и до середины ХХ века ученые не обращали никакого внимания на эти клетки, рассматривая их как вспомогательные элементы (клетки-спутники) нервной ткани. И только в последние 10–15 лет стало ясным, что глиальные клетки играют в процессах памяти, обучения и мышления не менее важную роль, чем нейроны.
Глию делят на «микроглию», «макроглию» и «радиальную глию». Макроглия представлена в ЦНС астроцитами и олигодендроцитами, а в периферической нервной системе — Шванновскими клетками. Еще один тип клеток макроглии — радиальная глия — играет крайне важную роль в формировании нервной ткани на разных стадиях эмбриогенеза и прижизненного нейрогенеза. Микроглия представлена особой популяцией клеток-фагоцитов нервной системы.
Глиальные клетки одного типа связаны между собой щелевыми синапсами и образуют единую электрически однородную сеть («синцитий»).
Макроглия играет важную роль в поддержании гомеостаза экстраклеточной среды. В этой связи мембраны глиальных клеток так же, как и мембраны нейронов, содержат ионные каналы, рецепторы нейромедиаторов, ионные насосы и транспортеры аминокислот. В мембранах астроцитов и Шванновских клеток имеются калиевые и хлорные ионные каналы, а также потенци- ал-зависимые натриевые и кальциевые каналы. Мембранный потенциал клеток макроглии полностью определяется потенциалом электрохимического равновесия для ионов калия. Глиальные клетки включены в процесс перераспределения ионов калия во внеклеточной среде, образуя в нервной системе «пространственный буфер экстраклеточной концентрации калия». В мембранах клеток глии обнаружены ионные помпы для трансмембранного транспорта ионов натрия и калия, бикарбоната и протонов. В мембранах астроцитов, олигодендроцитов и Шванновских клеток широко представлены рецепторы и транспортеры для
433

таких нейромедиаторов, как глутамат, ГАМК, глицин, ацетилхолин и норадреналин.
Функции глии не ограничиваются, как полагали до недавнего времени, снабжением нейронов питательными веществами (лактат, кислород, глюкоза). Олигодендроциты и Шванновские клетки осуществляют миелинизацию и ремиелинизацию (при повреждениях) аксонов нейронов в ЦНС и периферической нервной системе соответственно. Глия является важной составной частью гематоэнцефалического барьера. Глиальные клетки включены во все процессы мозга, требующие пласти- ческих перестроек нейронных сетей: эмбриогенез и прижизненный нейрогенез, формирование следов памяти, регенерация поврежденной нервной ткани.
14.1.Морфология глии
Â1846 г. немецкий врач и ученый Р. Вирхов1 ввел термин «глиальные клетки», или «глия» (от англ. «glue» — клей), для обозначе- ния открытой им своеобразной разновидности соединительной ткани, расположенной между нервными клетками и как бы соединяющей («склеивающей») их между собой. В 1883 г. знаменитый итальянский гистолог К.Гольджи обратил внимание на то, что глия отличается от обычной соединительной ткани морфологически и химически и имеет иное эмбриональное происхождение. По этой причине для обозначе- ния этой специфической соединительной ткани он предложил широко используемый в настоящее время термин «нейроглия» (синонимы:
клетки-сателлиты, клетки-спутники). Клетки глии способны делиться и размножаться во взрослом организме, что является наиболее частым источником образования опухолей мозга и «глиальных рубцов» в процессе регенерации пораженной нервной ткани.
Современные исследования демонстрируют принципиальное сходство в строении и свойствах глиальных клеток беспозвоночных (пиявка) и позвоночных (земноводные, млекопитающие) животных (Николлс
ñсоавт., 2003).
1Рудольф Вирхов (1821–1902) (íåì. Rudolf Ludwig Karl Virchow) — немецкий ученый и политический деятель, врач, патологоанатом, гистолог, физиолог, активно разрабатывавший основы клеточной теории в биологии и клеточ- ной патологии в медицине.
434
14.1.1.Типы глиальных клеток и их формирование
âэмбриогенезе
Общепринято делить «ãëèþ» íà «микроглию» è «макроглию», но некоторые исследователи относят к глии и «эпендимиальные клетки». Глиальных клеток всех типов примерно в 9–10 раз больше, чем нейронов. Это составляет около половины объема, занимаемого всей нервной системой (рис. 14.1; Филдз, 2004).
Ðèñ. 14.1. Глиальных клеток разного типа (на рисунке выделены красным) в нервной системе примерно в 9–10 раз больше, чем нейронов (выделены синим) (по: Филдз, 2004).
Различные морфологические типы глиальных клеток были идентифицированы с помощью методов световой флуоресцентной микроскопии и иммуногистохимии.
435
Макроглия включает «астроциты», «олигодендроциты», «Шванновские клетки» и «клетки радиальной глии» (рис. 14.2–14.5).
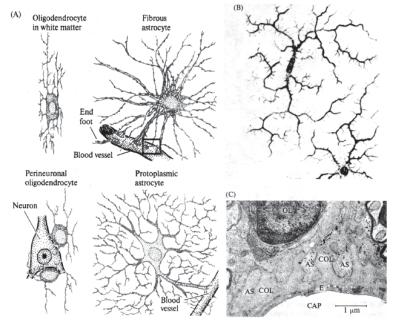
Ðèñ. 14.2. Типы нейроглиальных клеток в мозге млекопитающих (по: Николлс с соавт., 2003). (А) Олигодендроциты (oligodendrocytes), расположенные в белом веществе (in white matter) и вблизи тел нейронов (perineuronal); фиброзные (fibrous) и протоплазматические (protoplasmatic) астроциты. (B) Микроглиальные клетки. (С) Микрофотография рядом расположенных в оптическом тракте крысы кровеносного капилляра, фиброзного астроцита (AS) и олигодендроцита, сделанная с помощью электронного микроскопа. Обозначения: CAP
— просвет капилляра, E — эндотелиальные клетки внутренней стенки сосуда; COL — коллагеновые волокна, заполняющие пространство между между отростками AS и Е; OL — часть ядра олигодендроцита.
Астроциты контактируют, с одной стороны, с кровеносными капиллярами и, с другой, — с нейронами. Они делятся на два класса: фиброзные астроциты и протоплазматические астроциты (рис. 14.2, А). «Фиброзные астроциты» содержат большое количество филаментов и расположены преимущественно среди миелинизированных волокон. «Протоплазматические астроциты», содержащие меньше фиброзного материала, преобладают в области сомы, дендритов и синапсов нейронов.
436
Олигодендроциты (клетки овальной формы) сконцентрированы в центральной нервной системе (ЦНС), где они формируют миелиновую оболочку вокруг крупных аксонов (рис. 14.3–14.4). В нервах и ганглиях периферической нервной системы аналогами олигодендроцитов являются Шванновские клетки, открытые немецким физиологом и гистологом Т. Шванном в 1838 г. (рис. 14.5). Эти клетки формируют миелин вокруг крупных аксонов с высокой скоростью проведения возбуждения. Олигодендроциты и Шванновские клетки имеют разное происхождение в эмбриогенезе: первые образуются из клеток-предше- ственниц, выстилающих мозговую трубку (эпендимы), а вторые — из нервного гребня. Клетки радиальной глии с помощью своих отростков, натянутых, как «струны», образуют своеобразные пути (тракты), вдоль которых развивающиеся в процессе нейрогенеза нервные клетки мигрируют к местам своего назначения (рис. 14.18). Считается, что в ЦНС взрослых животных аналогами радиальной глии являются клетки Бергмана в мозжечке и Мюллеровские клетки в сетчатке глаза.
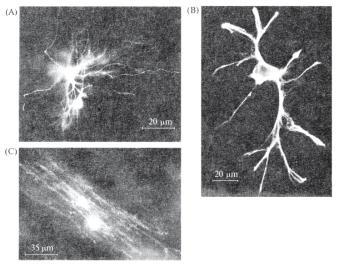
Ðèñ. 14.3. Клетки глии оптического нерва крысы, прокрашенные с помощью методов флуоресцентной микроскопии и иммунной гистохимии (по: Butt, Ransom, 1993 (A, C); Newman, 1985 (B)). (А) Флуоресцентный краситель, введенный в
астроцит оптического нерва, прокрасил также соседний астроцит, куда проник через щелевые (электрические) синапсы между глиальными клетками. (В) Фиброзный астроцит, окрашенный антителами к глиальному протеину GFAP. (C) Флуоресцирующий олигодендроцит, заполненный Procion Yellow.
437
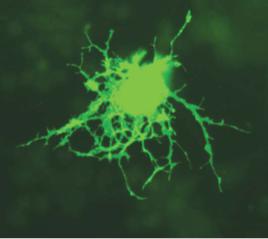
Ðèñ. 14.4. Олигоденроцит мыши, заполненный зеленым флуоресцирующим протеином GFP (Green Fluorescent Protein) (по: Broeke, 2008).
Микроглия. Микроглиальная ткань образована особой популяцией клеток-фагоцитов нервной системы. По строению и свойствам клетки микроглии напоминают макрофаги крови, от которых они, возмож-
но, и происходят (Brown, Perry, 1998).
Эпендимиальная ткань выстилает внутреннюю поверхность мозга (желудочков) и классифицируется как глиальная, так как является источником глиальных клеток в процессах эмбриогенеза и прижизненного нейрогенеза.
14.1.2. Синцитиальные объединения глиальных клеток
Мембраны соседних глиальных клеток плотно прилегают друг к другу и отделены узким экстраклеточным пространством в 2-3 нм, пронизанным тонкими протоплазматическими выростами, которые соединяют смежные локусы мембран двух клеток (рис. 14.6).
Протоплазматические «мостики», образованные скоплениями белковых молекул (коннексонов), позволяют осуществлять обмен ионами и мелкими молекулами между цитоплазмами смежных клеток. Такие плотные контакты называются «щелевыми соединениями (gap junctions)», или «электрическими (эфаптическими) синапсами». Они обладают низким сопротивлением и часто встречаются в нервной и других тканях организма (сетчатка глаза, эпителиальные клетки, гладкомышечные клетки сердечной мышцы). Таким образом, глиальные клетки одного
438
типа связаны между собой щелевыми синапсами и образуют единую электрически однородную сеть, так называемый «синцитий». Синцитиальные объединения глиальных клеток играют важную роль в обеспе- чении процессов межнейронных взаимодействий.
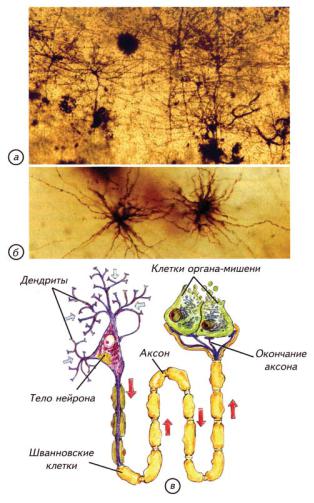
Ðèñ. 14.5. Глиальные клетки и нейроны человека, окрашенные по методу Гольджи (импрегнация серебром) (по: Савельев, 2005). (а) Нейроны коры головного мозга человека. (б) Глиальные клетки из мозга человека. (в) Нейроны и глия (Шванновские клетки) периферической нервной системы.
439