

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfприводит к возникновению последовательных залпов высокочастотных потенциалов действия без адаптации спайков по частоте. Многие из этих нейронов сохраняют без изменения паттерны разрядов и привязку к тета-ритму в присутствии антагониста мускарина. 3. Клетки с ПД большой длительности и выраженной медленной следовой гиперполяризацией имели низкую частоту разрядов и обычно только 1-3 спайка на тета-цикл, привязаны в основном к позитивному пику тета-волны зубчастой фасции. Гиперполяризационный ток вызываает прекращение генерации спайков и ярко выраженную адаптацию к его действию. ПД и мембранные свойства клеток этого типа совпадают с теми, которые были описаны для холинергических септальных нейронов. Тетапривязанная активность этих клеток уничтожалась мускариновыми антагонистами. Фазическое торможение холинергических MS/DB нейронов ГАБАергическими MS/DB нейронами сопровождалось эффектом отдачи. Предполагается, что это и является механизмом, который дает вклад в вовлечение MS/DB нейронной популяции в синхронизированные ритмические берсты активности, которые лежат в основе развития
гиппокампального тета-ритма (Brazhnik, Fox, 1997).
Пейсмекеры и синаптическая пластичность. Среди различ- ных областей мозга гиппокамп и неокортекс демонстрируют высокую чувствительность к индукции синаптической пластичности. В работе Yoshimura (2005) внимание фокусируется на этих кортикальных областях и исследуется действие кофеина на пластические изменения структуры и функции от синапса до кортикальных систем. Кофеин может влиять на функции мозга через повышение уровня внутриклеточного кальция. Обсуждая способы реорганизации нейронных систем, автор приходит к идее о том, что для этого может понадобиться синхронизация синаптической активности между кластерами нейронов. В этой связи между синаптической пластичностью и нейронными осцилляциями могут существовать весьма сильные зависимости. Ясно, что нача- ло осцилляций требует запускающего входа, а осцилляция обеспечит распространение нескольких колебаний. Индукция осцилляций требует низкочастотной активности от входных волокон в соединении с аппликацией кофеина. Отметим, что запуск осцилляций требует и активации NMDA-рецепторов, и высвобождения внутриклеточного кальция из внутренних кальциевых хранилищ. Предполагается наличие осциллирующих нейронов в зрительной коре. В других исследованиях показано, что осцилляторы могут быть и за пределами зрительной коры. Ретросплениальная кора занимает критическую позицию между зри-
390
тельной корой и гиппокампом. В ретросплениальной коре сциллятор присутствует в гранулярной ретросплениальной коре. Активация осцилляторов, находящихся во вторичной зрительной коре и в ретросплениуме, при аппликации кофеина открывает функциональные связи от первичной зрительной коры к постсубикулуму. Поэтому в присутствии кофеина осциллятор с локальной синхронизацией может индуцировать пространственно обширную синаптическую пластичность от зрительной коры к гиппокампальной формации. Это исследование инициировано «осцилляторно-зависимой» гипотезой пластичности (Yoshimura, 2005).
13.7. Пейсмекеры мозжечка
Клинические данные, полученные в исследованиях человека, и экспериментальные результаты на животных демонстрируют участие мозжечка в координации баллистических движений и сопровождающих их позных изменениях. Мозжечек участвует в кодировании порядка и длительности сокращения различных протагонистических мышц, опосредующих одно и то же движение. В обычной жизни это автомати- ческие, неосознаваемые действия. Последние исследования показали, что неоцеребеллум человека (латеральное полушарие и зубчатое ядро) играет важную роль в регуляции некоторых неокортикальных когнитивных функций. Этот новый функциональный аспект активности мозжечка интерферирует с результатами, полученными в трех разных ветвях исследований: нейроанатомическими данными, показывающими существование (иногда реципрокного) проводящего пути между неоцеребеллумом и ассоциативной и лимбической областями у приматов, нейрофизиологическими сведениями, демонстрирующими ухудшение чисто когнитивных функций у пациентов с нарушениями функции мозжечка, и данными магнито-резонансного сканирования на здовровых людях во время выполнения немоторных задач. Таким образом, церебеллум может участвовать в регуляции сенсорных, процедурных, лингвистических и эмоциональных активностей. Это означает, что операции на мозжечке могут сопровождаться когнитивными и аффективными синдромами в зависимости от важности и локализации операции (Habas, 2001). fMRI позволяет сравнить гемодинамические ответы, которые вызываются разными причинами при выполнении заданий, связанных с воспоминанием. Таким образом были идентифицированы те области мозга, которые делают вклад в успешное припоминание, включая
391
префронтальную кору и медиальную височную кору. Хотя участие мозжечка в вербальной рабочей памяти хорошо подтверждено, вклад мозжечка в эпизодическое долговременное кодирование было описано лишь вскользь, а механизмы, лежащие в основе памяти мозжечка, неясны (Fliessbach et al., 2007).
Общеизвестна удивительная однородность и регулярность структуры мозжечка, что привлекает к его исследованиям пристальное внимание физиологов, морфологов и теоретиков. Благодаря этому, детали нейронной организации мозжечка изучены на структурном и функциональном уровнях, по-видимому, лучше, чем характеристики других отделов мозга.
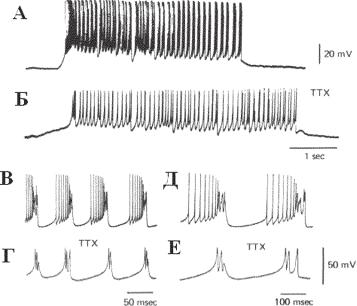
Ðèñ. 13.27. Спонтанные разряды клеток Пуркинье. А — спонтанная деполяризация, продолжающаяся 4 с, наблюдалась в отсутствии стимуляции. Б — добавление ТТХ вызывает блокаду соматического спайка. В–Д — спайковые разряды, состоящие из быстрого спайка и дендритного спайкового залпа. Г и Е — ответы после добавления в раствор ТТХ. Заметно, что спонтанные залпы спайков не изменяются, асоматические спайки блокируются (по R. LLINAS AND M. SUGIMORI Electrophysiological Properties Of In Vitro Purkinje Cell Dendrites In Mammalian Cerebellar Slices J. Physiol. (1980), 305, pp. 197-213).
392
Центральным элементом в мозжечке является клетка Пуркинье. Каждая клетка получает чрезвычайно много (до 500 тыс.) синапсов от аксонов зернистых клеток, формирующих ПВ. Помимо входов от большого числа зернистых клеток, на каждую КП через сложный синапс оказывает мощное воздействие одиночное афферентное нервное волокно — лианное волокно. Активность клеток Пуркинье чрезвычайно разнообразна: от длительных пауз и высокочастотных пачек импульсов до своеобразных относительно медленных высокоамплитудных потенциалов, растянутых нередко на десятки миллисекунд. Это так называемые сложные импульсы, возникающие со средней частотой 1 раз в секунду (рис. 13.27).
Нейроны мозжечка привлекли внимание исследователей необычайным разнообразием паттернов электрической активности. В работе Llinas and Sugimori (1980) регистрировалась электрическая активность клеток Пуркинье на срезах мозжечка морской свинки. Методом внутриклеточ- ного отведения исследовалась активность сомы, антидромное распространение импульса и электрическая и синаптическая чувствительность. Прямая электрическая стимуляция вызывала быстрые повторяющиеся разряды сомы, когда достигался порог генерации спайков. Минимальная частота разрядов около 30 сп./сек. При увеличении силы деполяризационного стимула наблюдался второй тип электрических ответов — залпы спайковой активности, которые часто сопровождались инактивацией соматических спайков. При аппликации тетродотоксина или удалении натрия из омывающего физраствора соматические спайки уничтожались, а залповая активность сохранялась. Однако блокирование кальциевой проводимости кобальтом, кадмием, марганцем, D600 или замещением кальция магнием в физрастворе убирали генерацию залповой активности. Если Са2+ блокировали или удаляли из раствора, не блокируя Na, то можно было наблюдать два типа натрий-чувствительных процессов: соматические спайковые разряды и медленные деполяризационные ответы, которые развивались по типу «все-или-ничего». Они достигали -15 мВ и могли продолжаться в течение нескольких секунд. Эти потенциалы блокировались ТТХ или исчезали в безнатриевом растворе. Спонтанная активность клеток Пуркинье характеризовалась соматическими спайками и залповыми разрядами. При добавлении ТТХ основная активность и ее частота оставались сохранными, демонстрируя зависимость от присутствия Са2+.
У клеток Пуркинье в ответ на стимуляцию восходящих волокон возникают как высокочастотные одиночные спайки, так и сложные ком-
393
плексные — залповые процессы. Было найдено, что натриевые спайки распространяются вплоть до частоты 260 Гц, а комплексные спайки проводятся не полностью, а только первый и последний спайк. Гиперполяризация повышает эффективность проведения, не влияя на общее число аксонных спайков, а деполяризация может уничтожить проведение сложного комплекса спайков (залпа). Эти данные показывают, что сложные комплексы не всегда проводятся по аксонам к нижележащим синапсам, а распространение ответов по восходящему волокну может модулироваться фоновой активностью (Monsivais et al., 2005). Хорошо известно, что входы клеток Пуркинье от восходящих волокон могут оказывать контролирующее влияние на фоновые простые спайки этих клеток, а повторная стимуляции восходящих волокон приводит к снижению спайковой активности. Удаление или инактивация этих волокон сопровождается увеличением спайковой активности клеток Пуркинье. В опытах Cerminara and Rawson (2004) показано, что если применить инактивацию клеток Пуркинье посредством влияния на входы от восходящих волокон вместе с веществом лигнокаином или микрохирургией верхних олив, то соматические разряды КП становятся значи- тельно более регулярными и достигают по частоте высокого уровня. Полученные результаты позволяют сделать вывод о том, что восходящие волокна оказывают на клетки Пуркинье тормозное действие. Тормозные входы модулируют работу осцилляторного механизма КП.
Хотя считается, что клетки Гольджи — основной тормозный интернейрон гранулярного слоя мозжечка — играют центральную роль в функционировании церебеллярной сети, их возбудительные свойства остаются неисследованными. Клетки Гольджи разряжаются ритмически in vivo и в срезах мозжечка, но непонятно, всегда ли эта активность связана с пейсмекерным ионным механизмом. В работе Forti et al. (2006), выполненной на срезах мозга трехнедельной крысы, регистрировалась активность нейронов. При комнатной температуре клетки Гольджи разряжались с частотой 1–10 Гц и 2–20 Гц. При температуре 35–37îС эта активность сохранялась в присутствии блокаторов быстрых синаптических рецепторов и рецепторов mGluR and GABAB, проявив себя в этих условиях как истинно пейсмекерные клетки. При использовании блокатора гиперполяризующего активационного тока развивалось падение частоты пейсмекера. Роль подпорогового натриевого тока не могли тестировать непосредственно, но наблюдалась четко выраженная TTX-чувствительность, не активирующая натриевый ток в подпороговой зоне. Когда исследовали ток реполяризации, то
394
обнаружили, что блокада кальций-зависимого калиевого тока снижает точность пейсмекера, не изменяя его частоты. Работа показала, что в пейсмекерную активность клеток Гольджи вносит вклад множество ионных механизмов, которые осуществляют тонкую регуляцию частоты генерации и точность ритма, влияя на циклическое торможение, вызываемое клетками Гольджи на гранулярном слое мозжечка.
13.8. Пейсмекеры верхних олив
Нейроны верхних олив (ВО) проявляют спонтанную осцилляторную активность, но значение этих осцилляций для формирования ответов этой системы не выяснено. Клетки Пуркинье оливо-церебеллярной системы генерируют ритмические и синхронные комплексные разряды. Предполагается, что этот феномен является результатом электротонического взаимодействия между клетками верхних олив и способностью этих клеток генерировать осцилляторную активность. То, что клетки верхних олив являются спонтанными осцилляторами, поддерживается данными нескольких направлений исследований: ВО-клет- ки, зарегистрированные на срезах мозга (в которых их афферентные входы существенно ограничиваются), проявляют подпороговые осцилляции мембранного потенциала (Llina´s and Yarom, 1981, 1986); ритми- ческая активность комплексными спайками может быть изменена при помощи инъекции апамина и харибдотоксина, влияющих на проводимость Са2+, ритмическая активность комплексных спайков не только сохраняется, но и действительно становится более выраженной после блокады глютаматэргических и ГАМКэргических входов к нейронам ВО у интактных животных. Относительно мало исследований, которые могли бы ответить на вопрос о том, как эти осцилляторные свойства содействуют взаимодействию оливоцеребеллярной системы с другими областями мозга. Большая часть их делала вывод: подпороговые осцилляции ВО нейронов могут вызывать их ответы на синаптическую активность в зависимости от фазы цикла, на которую попадает синаптическая активность. Сильный стимул запускает ВО цикл осцилляций и продуцирует сцепленные осцилляторы комплексных спайков или спайковый ответ нейронов ВО. Предполагается, что осцилляторные свойства ВО нейронов определяют, как именно будет взаимодействовать оливоцеребеллярная система с другими областями мозга, но отсутствуют соображения о том, как будут ВО нейроны взаимодействовать со входящими осциллирующими сигналами, которые вполне могут исходить из моторной коры (Lang, 2001). Действительно, проводящие пути
395
из двигательной коры обеспечивают значимые функциональные входы на ВО, потому что электрическая стимуляция двигательной коры охотно запускает комплексные спайки. Так, эти две системы формируют организацию, в которой осцилляторная активность одной системы (моторной коры) может управлять осцилляторной активностью другой системы (оливоцеребеллярной).
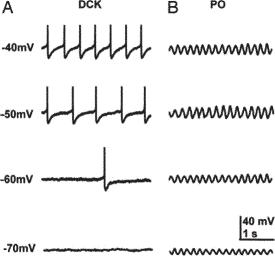
Ðèñ. 13.28. Повторные разряды нейронов мозжечка (дорзального конца Kooy) зависят от мембранного потенциала. А— регистрация активности при МП -70, -60, -50 и -40 мВ. Частота разрядов увеличивается при деполяризации. Каждый отдельный потенциал действия сопровождается большой, продолжительной следовой гиперполяризацией. Амплитуда ПД на этом рисунке обрезана, чтобы продемонстрировать гиперполяризационное последействие и динамику развития потенциала. Б — регистрация ПО нейрона показывает подпороговые осцилляции частотой около 5 Гц при таких же значениях МП, что и на нейрограммах А. DCK нейрон и ПО нейрон зарегистрированы на одном и том же срезе (по Proc Natl Acad Sci U S A. 2006 Oct 31;103(44):16550-5. Epub 2006 Oct 18. Somatomotor and oculomotor inferior olivary neurons have distinct electrophysiological phenotypes. Urbano FJ, Simpson JI, Llinás RR).
В экспериментах Marshall и Lang (2004) применялся метод множественной электродной регистрации, чтобы изучить оливоцеребеллярные ответы на активацию моторного кортико-оливарного пути у крысы. К двигательной коре применялся ряд электрических стимулов часто-
396
той от 4 до 30 Гц. Кривые соотношения «частота стимула — величина ответной реакции» показали, что наиболее общим свойством всех нейронов является унимодальность с максимумом около 9,5 Гц и бимодальность с пиками около 8,9 Гц и 15,1 Гц. Чтобы определить, действительно ли в основе осцилляторных свойств ВO нейронов лежат резонансные пики этих кривых, в область изучения нейронов ВO инъецировали токсины, которые ослабляют или усиливают спонтанные осцилляции ВO, оказывая соответствующее влияние на остроту резонансных пиков. Результаты опытов показали, что нейроны верхних олив отвечают на афферентные сигналы как нелинейные осцилляторы. Это означает, что ВО действует как нелинейный осциллятор, который предпочитает отвечать на частоты вокруг относительно узкого частотного диапазона около 10 Гц.
Электрофизиологические свойства нейронов верхних олив крысы дорзального конца и восходящего вентролатерального отростка сравнивались с нейронами основной оливы (ОО) (рис. 13.28). В то время как нейроны DCK/VLO вовлекаются в контроль движения глаз через проекцию их восходящих волокон в церебеллярный флоккулус (выступ на коре мозжечка), ВO нейроны контролируют конечности и движение пальцев через проекцию своих восходящих волокон к латеральной церебеллярной полусфере. В опытах in vitro при регистрации активности нейронов DCK/VLO выяснилось, что низкий порог кальциевого тока и подпороговые осцилляции отсутствуют у этих нейронов. Регистрация активности DCK нейронов, полученная при использовании изображения вольт-чувствительной краской, показала, что активность не ограничивается одним нейроном, а скорее кластерами нейронов DCK, которые могут быть активированы в унисон. Эти электрофизиологи- ческие эксперименты показали, что нейроны DCK/VLO имеют уникальные свойства, которые выводят их за пределы нейронов PO ядра. Эта находка показывает, что моторный контроль, исходящий из оливоцеребеллярной системы, фундаментально отличает ее от окуломотор-
ной и соматомоторной систем (Urbano et al., 2006).
Осцилляторные свойства нейронов верхних олив. Известно, что в опытах in vivo нейроны ВО осциллируют на частотах 5-10/ñ. с необыкновенной регулярностью. Взаимоотношение между проводимостями, описанными в работе Llinas and Y. Yarom (1981), должны выявить много неожиданных аспектов осцилляторного поведения нейронов ВО. Они должны быть способными запускать механизм генерации или при антидроной или при ортодромной активации, которая заставит
397
генерировать ADP-AHP последовательности. Осциллирующие кальциевые спайки развиваются на двух частотах, каждая из которых генерируется своим механизмом. Если клетка (особенно дендриты) частично деполяризована фоновой активностью, активация Na+ спайка может успешно генерировать разряды с дендритов, которые продуцируют ребаунд соматического кальциевого спайка. В других случаях, связанных с негативностью мембранного потенциала, Na+ не может активировать дендритный потенциал, но может запускать последовательность Са-К активаций на соме, которая, в свою очередь, приводит к сомати- ческому ребаунду. В таких случаях частота разрядов может быть до 15 спайков/сек. вместо обычных 5-8 спайков/сек. Модуляция уровня мембранного потенциала и дендритная возбудимость, по-видимому, оказываются существенным параметром для определения частоты ос-
цилляций этих нейронов (Llinas and Yarom, 1981).
Тормозные влияния от восходящих волокон. Хорошо известно, что входы клеток Пуркинье от восходящих волокон могут оказывать контролирующее влияние на фоновые простые спайки этих клеток, а повторная стимуляции восходящих волокон приводит к снижению спайковой активности. Удаление или инактивация этих волокон сопровождается увеличением спайковой активности клеток Пуркинье. В опытах Cerminara and Rawson (2004) показано, что если применить инактивацию клеток Пуркинье посредством влияния на входы от восходящих волокон вместе с веществом лигнокаином или микрохирургией верхних олив, то соматические разряды клеток Пуркинье становятся значительно более регулярными и достигают по частоте высокого уровня. Полученные результаты позволяют сделать вывод о том, что восходящие волокна оказывают на клетки Пуркинье тормозное действие. Тормозные входы модулируют работу осцилляторного механизма клеток Пуркинье.
13.9. Пейсмекеры гипоталамуса
Нейроны переднего преоптического ядра чувствуют и регулируют температуру тела. Хотя в противовес этому существует и иное мнение: вызываемая теплом деполяризация определяет нейронную термочувствительность. Поддерживая эту гипотезу, недавние исследования показали, что катион-чувствительные ионные каналы включают в свой состав основополагающий механизм нейронной термочувствительности. Кроме того, было показано в более ранних работах, что теплочувствительность нейронов переднего преоптического гипоталамуса свя-
398
зана с деполяризационным натриевым током, который чувствителен к тетродотоксину. Чтобы это проверить, провели опыты с регистрацией от срезов гипоталамуса мозга крысы. Сравнили термальные эффекты на термочувствительных нейронах, температурочувствительных и мол- чащих нейронах. Все три типа нейронов показывали небольшую деполяризацию при действии тепла и гиперполяризацию при действии холода. В опытах не обнаружили следов resting-ионных токов, которые являются уникальными для теплочувствительных нейронов. Это подтверждают те исследования, в которых предполагается, что нейронная термочувствительность находится под контролем токов, определяющих быстрые изменения мембранного потенциала между последовательными потенциалами действия (Zhao and Boulant, 2005).
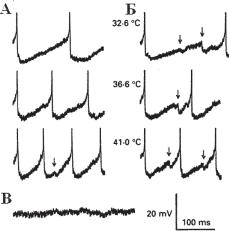
Ðèñ. 13.29. Термочувствительные нейроны гипоталамуса. А — активность преоптического тепло-чувствительного нейрона, зарегистрированного при трех разных температурах . Потенциал покоя примерно -60 мВ. Стрелки показывают приход ТПСП-подобной активности. Б — активность того же самого нейрона, но межспайковые интералы более длинные, и здесь представлено большее количе- ство ТПСП. В — активность при 41îС во время инъекции гиперполяризационного тока (по J Physiol. 1991; 440: 257-71, Curras MC, Kelso SR, Boulant J.A.).
В исследовании Curras, Kelso and Boulant (1991), выполненном методом внутриклеточной регистрации на нейронах гипоталамуса, показано, что примерно 30% нейронов являются чувствительными к теплу, 10% к холоду, 60% температурно нечувствительны. Авторы обнару-
399