

книги студ / color atlas of physiology 5th ed[1]. (a. despopoulos et al, thieme 2003)
.pdf4 |
|
Blood |
|
|
|
|
|
|
||
|
Composition and Function of Blood |
(e.g., heme) can be protected from breakdown |
||||||||
|
and renal excretion. The binding of small |
|||||||||
|
|
|
|
|
|
|||||
|
The blood volume of an adult correlates with |
molecules to plasma proteins reduces their |
||||||||
|
his or her (fat-free) body mass and amounts to |
osmotic efficacy. Many plasma proteins are in- |
||||||||
|
ca. 4–4.5 L in women (!) and 4.5–5 L in men of |
volved in blood clotting and fibrinolysis. |
||||||||
|
70 kg BW ("; !table). The functions of blood |
Serum forms when fibrinogen separates from |
||||||||
|
include the transport of various molecules (O2, |
plasma in the process of blood clotting. |
||||||||
|
CO2, nutrients, metabolites, vitamins, electro- |
The formationofbloodcells occursinthered |
||||||||
|
lytes, etc.), heat (regulation of body tempera- |
bone marrow of flat bone in adults and in the |
||||||||
|
ture) and transmission of signals (hormones) as |
spleen and liver of the fetus. Hematopoietic tis- |
||||||||
|
well as buffering and immune defense. The |
suescontainpluripotent stemcells which,with |
||||||||
|
blood consists of a fluid (plasma) formed el- |
the aid of hematopoietic growth factors (see |
||||||||
|
ements: Red blood cells (RBCs) transport O2 |
below), develop into myeloid, erythroid and |
||||||||
|
and play an important role in pH regulation. |
lymphoid precursor cells. Since pluripotent |
||||||||
|
White blood cells (WBCs) can be divided into |
stem cells are autoreproductive, their existence |
||||||||
|
neutrophilic, |
eosinophilic and basophilic |
is ensured throughout life. In lymphocyte |
|||||||
|
granulocytes, monocytes, and lymphocytes. |
development, lymphocytes arising from lym- |
||||||||
|
Neutrophils play a role in nonspecific immune |
phoid precursor cells first undergo special |
||||||||
|
defense, whereas monocytes and lymphocytes |
differentiation (in the thymus or bone marrow) |
||||||||
|
participate in specific immune responses. |
and are later formed in the spleen and lymph |
||||||||
|
Platelets (thrombocytes) are needed for he- |
nodes as well as in the bone marrow. All other |
||||||||
|
mostasis. Hematocrit (Hct) is the volume ratio |
precursor cells are produced by myelocytopoie- |
||||||||
|
of red cells to whole blood (!C and Table). |
sis, that is, the entire process of proliferation, |
||||||||
|
Plasma is the fluid portion of the blood in |
maturation, and release into the bloodstream |
||||||||
|
which electrolytes, nutrients, metabolites, vi- |
occurs in the bone marrow. Two hormones, er- |
||||||||
|
tamins, hormones, gases, and proteins are dis- |
ythropoietin and thrombopoietin, are involved |
||||||||
|
solved. |
|
|
|
in myelopoiesis. Thrombopoietin |
(formed |
||||
|
|
Plasma proteins (!Table) are involved in |
mainly in the liver) promotes the maturation |
|||||||
|
humoral immune defense and maintain on- |
and development of megakaryocytes from |
||||||||
|
cotic pressure, which helps to keep the blood |
which the platelets are split off. A number of |
||||||||
|
volume constant. By binding to plasma pro- |
othergrowthfactorsaffectbloodcellformation |
||||||||
|
teins, compounds insoluble in water can be |
in bone marrow via paracrine mechanisms. |
||||||||
|
transported in blood, and many substances |
Erythropoietin promotes the maturation and |
||||||||
|
|
|
|
|
|
proliferation of red blood cells. It is secreted by |
||||
|
Blood volume in liters relative to body weight (BW) |
|||||||||
|
the liver in the fetus, and chiefly by the kidney |
|||||||||
|
" 0.041!BW (kg) + 1.53, ! 0.047 !BW (kg) + 0.86 |
(ca. 90%) in postnatal life. In response to an oxy- |
||||||||
|
Hematocrit (cell volume/ blood volume): |
|||||||||
|
gen deficiency (due to high altitudes, hemoly- |
|||||||||
|
" 0.40–0.54 |
Females: 0.37–0.47 |
sis, etc.; !A), erythropoietin secretion in- |
|||||||
|
Erythrocytes (1012/L of blood = 106/ µL of blood): |
|||||||||
|
creases, larger numbers of red blood cells are |
|||||||||
|
" 4.6–5.9 |
! 4.2–5.4 |
produced, and the fraction of reticulocytes |
|||||||
|
Hemoglobin (g/L of blood): |
|||||||||
|
(young erythrocytes) in the blood rises. The life |
|||||||||
|
"140–180 |
! 120–160 |
span of a red blood cell is around 120 days. Red |
|||||||
|
MCH, MCV, MCHC—mean corpuscular (MC), hemo- |
|||||||||
|
blood cells regularly exit from arterioles in the |
|||||||||
|
globin (Hb), MC volume, MC Hb concentration !C |
splenic pulp and travel through small pores to |
||||||||
|
Leukocytes (109/L of blood = 103/ µL of blood): |
|||||||||
|
enter the splenic sinus (!B), where old red |
|||||||||
|
3–11 (64% granulocytes, 31% lymphocytes, |
blood cells are sorted out |
and |
destroyed |
||||||
|
6% monocytes) |
|||||||||
|
(hemolysis). Macrophages in the spleen, liver, |
|||||||||
|
Platelets (109/L of blood = 103/ µL of blood): |
|||||||||
|
bone marrow, etc. engulf and break down the |
|||||||||
|
" 170–360 |
|
!180–400 |
|||||||
88 |
|
cell fragments. Heme, |
the |
iron-containing |
||||||
Plasma proteins (g/L of serum): |
||||||||||
group of hemoglobin |
(Hb) |
released during |
||||||||
|
66–85 (including 55–64% albumin) |
|||||||||
|
hemolysis, is broken |
down into |
bilirubin |
|||||||
|
|
|
|
|
|
|||||
(!p. 250), and the iron is recycled (!p. 90).
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

A. Regulation of RBC production |
B. Life cycle of red blood cells |
|
|
||
1 Hypoxia |
|
Bone marrow |
|
|
|
|
|
|
|
||
PO2 |
High altitude, etc. |
|
RBC formation |
|
|
|
|
|
|
Bloodof |
|
|
PO2 |
|
|
|
|
Kidney |
|
Life span: |
|
|
Function |
|
Erythrocytes |
120 days |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
Erythropoietin |
|
|
|
|
|
|
|
Blood |
|
|
and |
|
|
|
|
|
|
PO2 |
Hemolysis |
down |
|
|
Composition |
|
Bone marrow |
|
|
|
|
2 Hemolysis |
|
Break- |
|
|
|
|
|
|
|
|
|
|
|
Spleen |
|
“Still good” |
4.1 |
|
|
|
|
||
|
PO2 |
|
Test |
|
|
|
|
|
Plate |
||
|
|
“Too old” |
|
|
|
|
|
|
|
|
|
|
|
Phagocytosis |
|
Spenic |
|
|
|
|
pulp |
|
|
|
|
by |
|
|
|
|
|
Pulpal |
|
|
|
Erythropoietin |
|
macrophages in: |
|
|
|
|
|
Bone marrow |
arteriole |
|
|
|
|
|
|
|
|
|
|
Lymph nodes |
|
Sinus |
|
|
|
Spleen |
|
|
|
|
|
Liver, etc. |
|
|
|
C. Erythrocyte parameters MCH, MCV and MCHC
|
Centrifugation |
|
|
|
|
Blood sample |
a |
MCH (mean Hb mass/RBC) |
|||
|
|||||
|
|
||||
|
b |
= |
Hb conc. |
(g/RBC) |
Normal: |
|
|
|
red cell count |
|
|
|
|
|
|
|
27–32pg |
|
Hematocrit (Hct)=b/a |
|
|
|
|
|
(LRBC/LBlood) |
MCV (mean volume of one RBC) |
|||
|
|
||||
|
|
|
Hct |
|
|
|
Hemoglobin concentration |
= red cell count(L/RBC) |
Normal: |
||
Red cell count (RCC) |
(g/LBLOOD) |
|
|
|
80–100fl |
|
|
|
|
||
|
|
|
|
|
|
(quantity/LBLOOD) |
|
|
|
|
|
|
|
MCHC (mean Hb conc. in RBCs) |
|||
|
|
= |
Hb conc. |
(g/LRBC) |
Normal: |
|
|
|
Hct |
|
|
|
|
|
|
|
320–360g/L |
89
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

4 Blood
90
Iron Metabolism and Erythropoiesis
Roughly 2/3 of the body’s iron pool (ca. 2 g in women and 5 g in men) is bound to hemoglobin (Hb). About 1/4 exists as stored iron (ferritin, hemosiderin), the rest as functional iron (myoglobin, iron-containing enzymes). Iron losses from the body amount to about 1 mg/day in men and up to 2 mg/day in women due to menstruation, birth, and pregnancy. Iron absorption occurs mainly in the duodenum and varies according to need. The absorption of iron supplied by the diet usually amounts to about 3 to 15% in healthy individuals, but can increase to over 25% in individuals with iron deficiency (!A1). A minimum daily iron intake of at least 10–20 mg/day is therefore recommended (women ! children ! men).
Iron absorption (!A2). Fe(II) supplied by the diet (hemoglobin, myoglobin found chiefly in meat and fish) is absorbed relatively efficiently as a heme-Fe(II) upon protein cleavage. With the aid of heme oxygenase, Fe in mucosal cells cleaves from heme and oxidizes to Fe(III). The triferric form either remains in the mucosa as a ferritin-Fe(III) complex and returns to the lumen during cell turnover or enters the bloodstream. Non-heme-Fe can only be absorbed as Fe2+. Therefore, non-heme Fe(III) must first be reduced to Fe2+ by ferrireductase (FR; !A2) and ascorbate on the surface of the luminal mucosa (!A2). Fe2+ is probably absorbed through secondary active transport via an Fe2+-H+ symport carrier (DCT1) (competition with Mn2+, Co2+, Cd2+, etc.). A low chymous pH is important since it (a) increases the H+ gradient that drives Fe2+ via DCT1 into the cell and (b) frees dietary iron from complexes. The absorption of iron into the bloodstream is regulated by the intestinal mucosa. When an iron deficiency exists, aconitase (an iron-regu- lating protein) in the cytosol binds with fer- ritin-mRNA, thereby inhibiting mucosal ferritin translation. As a result, larger quantities of absorbed Fe(II) can enter the bloodstream. Fe(II) in the blood is oxidized to Fe(III) by ceruloplasmin (and copper). It then binds to apotransferrin, a protein responsible for iron transport in plasma (!A2, 3). Transferrin
(= apotransferrin loaded with 2 Fe(III)), is taken up by endocytosis into erythroblasts and cells of the liver, placenta, etc. with the aid of
transferrin receptors. Once iron has been released to the target cells, apotransferrin again becomes available for uptake of iron from the intestine and macrophages (see below).
Iron storage and recycling (!A3). Ferritin, one of the chief forms in which iron is stored in the body, occurs mainly in the intestinal mucosa, liver, bone marrow, red blood cells, and plasma. It contains binding pockets for up to 4500 Fe3+ ions and provides rapidly available stores of iron (ca. 600 mg), whereas iron mobilization from hemosiderin is much slower (250 mg Fe in macrophages of the liver and bone marrow). Hb-Fe and heme-Fe released from malformed erythroblasts (so-called inefficient erythropoiesis) and hemolyzed red blood cells bind to haptoglobin and hemopexin, respectively. They are then engulfed by macrophages in the bone marrow or in the liver and spleen, respectively, resulting in 97% iron recycling (!A3).
An iron deficiency inhibits Hb synthesis, leading to hypochromic microcytic anemia: MCH "26 pg, MCV "70 fL, Hb "110 g/L. The primary causes are:
blood loss (most common cause); 0.5 mg Fe are lost with each mL of blood;
insufficient iron intake or absorption;
increased iron requirement due to growth, pregnancy, breast-feeding, etc.;
decreased iron recycling (due to chronic infection);
apotransferrin defect (rare cause).
Iron overload most commonly damages the liver, pancreas and myocardium (hemochromatosis). If the iron supply bypasses the intestinal tract (iron injection), the transferrin capacity can be exceeded and the resulting quantities of free iron can induce iron poisoning.
B12 vitamin (cobalamins) and folic acid are also required for erythropoiesis (!B). Deficiencies lead to hyperchromic anemia (decreased RCC, increased MCH). The main causes are lack of intrinsic factor (required for cobalamin resorption) and decreased folic acid absorption due to malabsorption (see also p. 260) or an extremely unbalanced diet. Because of the large stores available, decreased cobalamin absorption does not lead to symptoms of deficiency until many years later, whereas folic acid deficiency leads to symptoms within a few months.
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

A.Iron intake and metabolism
1Iron intake
Normal Fe intake:  10–20 mg/day
10–20 mg/day
5–10 mg/day
Fe
Fe absorption:
3–15% of HCI Fe intake
Fe |
Stomach |
Liver
Non-absorbed Fe in feces:
Normally 85–97% of intake
3 Fe storage and Fe recycling
Bone |
Liver |
marrow |
|
|
Ferritin |
|
Fe |
|
Hemo- |
|
siderin |
|
Fe stores |
2 Fe absorption
Lumen |
2+ |
|
III |
|
Mucosal cells |
Blood -Apo transferrin |
|
|
|
Heme- |
(duodenum) |
|
|||
|
|
FeII |
|
|
Heme |
|
|
|
|
|
|
|
|
|
|
Fe |
|
Fe |
|
FR |
|
Mucosal |
|
|
|
|
|
Fe |
transferrin |
|
|
|
|
Fe2+ |
|
|
|
||
|
|
|
|
|
|
||
|
|
|
|
|
|
Ferritin |
|
|
|
|
H+ |
|
FeIII |
Lyso- |
Transferrin |
|
|
|
|
|
|
some |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cell |
FeIII FeIII |
|
|
|
|
|
FeIII |
turnover |
|
|
|
|
|
|
|
|
|
Transferrin
 Fe
Fe
Systemic |
|
|
|
blood |
|
|
|
Hemo- |
Heme |
Fe |
Ferritin |
pexin |
|
|
|
Hapto- |
Hb |
|
Hemo- |
globin |
|
siderin |
|
 Erythrocytes
Erythrocytes
Already in bone marrow |
Macrophages |
|
in spleen, liver and |
|
bone marrow (extravascular) |
B. Folic acid and vitamin B12 (cobalamins)
Folic acid |
Other organs |
|
||
0.05mg/day |
|
|
|
|
Vitamin B12 |
|
|
|
|
0.001mg/day |
Stores |
|
|
|
|
|
1 mg |
Methyl- |
|
|
|
|
||
|
|
|
cobalamin |
|
Intrinsic |
|
|
|
|
|
7mg |
|
|
|
factor |
|
N5-tetra- |
||
|
|
|||
|
|
|
hydrofolate |
|
|
Liver |
|
|
|
Stomach |
|
|
Erythropoiesis |
|
|
|
|
||
|
|
Erythrocytes |
Erythroblast |
|
Ileum
NADP
 NADPH +H+
NADPH +H+
Dihydrofolate reductase
|
|
|
7,8- |
|
|
Folate |
|
dihydro- |
|
|
|
folate |
||
regeneration |
||||
|
||||
Tetrahydro- |
Thymidylate |
|
||
folate |
synthase |
|
|
|
|
|
|
||
Deoxy- |
|
Deoxy- |
||
uridylate |
|
thymidylate |
||
DNA synthesis
Bone marrow
Plate 4.2 Iron Metabolism and Erythropoiesis
91
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

Flow Properties of Blood
|
The viscosity (η) of blood is higher than that of |
||||
|
plasma due to its erythrocyte (RBC) content. |
||||
|
Viscosity (η) = 1/fluidity = |
shearing force |
|||
|
(τ)/shearing action (γ) [Pa · s]. The viscosity of |
||||
|
blood rises with increasing hematocrit and |
||||
|
decreasing flow velocity. Erythrocytes lack the |
||||
|
major organelles and, therefore, are highly de- |
||||
|
formable. Because of the low viscosity of their |
||||
|
contents, the liquid film-like characteristics of |
||||
|
their membrane, and their high surface/ |
||||
|
volume ratio, the blood behaves more like an |
||||
|
emulsion than a cell suspension, especially |
||||
|
when it flows rapidly. The viscosity of flowing |
||||
|
blood (ηblood) passing through small arteries (! |
||||
|
20 µm) is about 4 relative units (RU). This is |
||||
|
twice as high as the viscosity of plasma (ηplasma |
||||
|
= 2 RU; water: 1 RU = 0.7 mPa · s at 37 "C). |
||||
Blood |
Because they are highly deformable, normal |
||||
RBCs normally have no problem passing |
|||||
|
|||||
4 |
through capillaries or pores in the splenic ves- |
||||
sels (see p. 89 B), although their diameter (! |
|||||
|
|||||
|
#5 µm) is smaller than that of freely mobile |
||||
|
RBCs (7 µm). Although the slowness of flow in |
||||
|
small vessels causes the blood viscosity to in- |
||||
|
crease, this |
is partially compensated for |
|||
|
(ηblood!) by the passage of red cells in single |
||||
|
file through the center of small vessels (diame- |
||||
|
ter #300 µm) due to the Fåhraeus–Lindqvist |
||||
|
effect ("A). Blood viscosity is only slightly |
||||
|
higher than |
plasma viscosity in |
arterioles |
||
|
(!!7 µm), |
but rises again |
in |
capillaries |
|
|
(!!4 µm). A critical increase |
in blood viscos- |
|||
|
ity can occur a) if blood flow becomes too slug- |
||||
|
gish and/or b) if the fluidity of red cells |
||||
|
decreases due to hyperosmolality (resulting in |
||||
|
crenation), cell inclusion, hemoglobin malfor- |
||||
|
mation (e.g., sickle-cell anemia), changes in |
||||
|
the cell membrane (e.g., in old red cells), and so |
||||
|
forth. Under such circumstances, the RBCs un- |
||||
|
dergo aggregation (rouleaux formation), in- |
||||
|
creasing the blood viscosity tremendously (up |
||||
|
to 1000 RU). This can quickly lead to the cessa- |
||||
|
tion of blood flow in small vessels ("p. 218). |
||||
|
Plasma, Ion Distribution |
|
|
||
|
Plasma is obtained by preventing the blood |
||||
92 |
from clotting and extracting the formed el- |
||||
ements by |
centrifugation ("p. 89 C). High |
||||
|
|||||
molecular weight proteins ("B) as well as ions and non-charged substances with low molecular weights are dissolved in plasma. The sum of the concentrations of these particles yields a plasma osmolality of 290 mOsm/ kgH2O ("pp. 164, 377). The most abundant cation in plasma is Na+, and the most abundant anions are Cl– and HCO3–. Although plasma proteins carry a number of anionic net charges ("C), their osmotic efficacy is smaller because the number of particles, not the ionic valency, is the determining factor.
The fraction of proteins able to leave the blood vessels is small and varies from one organ to another. Capillaries in the liver, for example, are much more permeable to proteins than those in the brain. The composition of interstitial fluid therefore differs significantly from that of plasma, especially with respect to protein content ("C). A completely different composition is found in the cytosol, where K+ is the prevailing cation, and where phosphates, proteins and other organic anions comprise the major fraction of anions ("C). These fractions vary depending on cell type.
Sixty percent of all plasma protein ("B) is albumin (35–46 g/L). Albumin serves as a vehicle for a number of substances in the blood. They are the main cause of colloidal osmotic pressure or, rather, oncotic pressure ("pp. 208, 378), and they provide a protein reserve in times of protein deficiency. The α1, α2 and % globulins mainly serve to transport lipids (apolipoproteins), hemoglobin (haptoglobin), iron (apotransferrin), cortisol (transcortin), and cobalamins (transcobalamin). Most plasma factors for coagulation and fibrinolysis are also proteins. Most plasma immunoglobulins (Ig, "D) belong to the group of γ globulins and serve as defense proteins (antibodies). IgG, the most abundant immunoglobulin (7–15 g/L), can cross the placental barrier (maternofetal transmission; "D). Each Ig consists of two group-specific, heavy protein chains (IgG: γ chain, IgA: α chain, IgM: µ chain, IgD: δ chain, IgE: ε chain) and two light protein chains (λ or κ chain) linked by disulfide bonds to yield a characteristic Y-shaped configuration (see p. 95 A).
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.
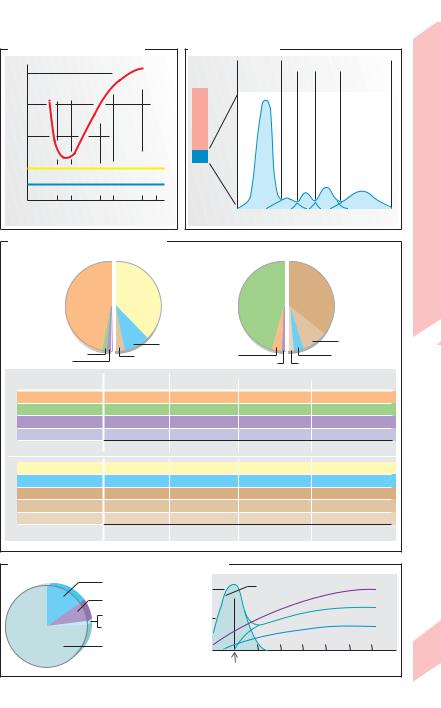
A. Fåhraeus-Lindqvist effect |
B. Plasmaproteins |
|
|
|
|
||||||||||
|
8 |
|
|
|
|
|
|
|
Albumin |
Globulins |
|
||||
|
|
|
|
|
Blood |
|
|
|
|
α1 |
α2 |
β |
γ |
|
|
units |
|
|
|
|
|
|
|
|
60% |
|
4% |
8% |
12% |
16% |
|
6 |
|
|
|
|
|
Plasma |
|
|
|
|
|
|
|
|
|
Viscosity in relative |
|
|
|
|
|
|
|
|
|
|
|
|
|
||
4 |
|
|
|
|
|
65–80 g/L Proteins (100%) |
|
|
|
|
|
of Blood |
|||
2 |
|
|
|
|
Plasma |
|
|
|
|
|
|||||
|
|
|
|
Water |
|
|
|
|
|
||||||
1 |
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Properties |
|
|
01 |
5 |
10 |
50 100 |
500 1000 |
|
|
Electrophoretic protein fractions |
|||||||
|
|
Vessel inside diameter (µm) |
|
|
|||||||||||
C. Ion composition of body fluids |
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
Flow |
|||||||
|
|
|
|
Interstitium |
|
|
|
|
Cytosol |
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cations |
|
Anions |
|
|
Cations |
|
|
|
Anions |
4.3 |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Na+ |
|
Cl– |
|
|
K+ |
|
Proteins– |
|
Plate |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
HCO3– |
|
|
|
|
|
|
|
Inorganic |
|
|
|
|
|
K+ |
|
|
|
|
|
|
|
|
phosphate |
|
|
|
2+ |
|
2+ |
|
Proteins,phosphates, |
Na+ |
|
2+ |
|
HCO3– |
|
||||
|
Ca ,Mg |
|
|
|
etc. |
|
|
2+ |
|
Misc. |
|
|
|||
|
|
|
|
|
|
|
|
Ca ,Mg |
|
|
|
||||
|
|
|
|
|
|
|
|
|
mval/L (mmol/L) |
|
|
|
|
||
|
Ion |
|
|
|
Plasma |
Serum |
|
Interstitium |
|
Cytosol |
|
||||
Cations |
Na+ |
|
|
|
|
142 |
153 |
|
145 |
|
|
ca. 12 |
|
||
K+ |
|
|
|
|
4.3 |
4.6 |
|
|
4.4 |
|
ca. 140 |
|
|||
Free Ca2+ |
|
|
2.6 (1.3*) |
2.8 (1.3) |
2.5 (1.5) |
|
<0,001 |
|
|||||||
Free Mg2+ |
|
|
1.0 (0.5**) |
1.0 (0.5) |
0.9 (0.45) |
|
|
1.6 |
|
||||||
|
|
|
|
|
|
||||||||||
|
|
|
Sum |
|
150 |
162 |
|
|
153 |
|
ca. 152 |
|
|||
|
Cl– |
|
|
|
|
104 |
112 |
|
|
117 |
|
|
ca. 3 |
|
|
Anions |
HCO3– |
|
|
|
|
24 |
36 |
|
|
27 |
|
|
10 |
|
|
Inorganic phosphate |
|
2 |
2.2 |
|
|
2.3 |
|
|
ca. 30 |
|
|||||
Proteins |
|
|
|
|
14 |
15 |
|
|
0.4 |
|
|
ca. 54 |
|
||
|
Misc. |
|
|
|
|
5.9 |
6.3 |
|
|
6.2 |
|
|
ca. 54 |
|
|
|
|
|
Sum |
|
150 |
162 |
|
|
153 |
|
ca. 152 |
|
|||
|
|
|
|
|
|
*) Total plasma Ca: 2.5mmol/L; **) Total plasma Mg: 0.9mmol/L |
|
||||||||
D. Concentrations of immunglobulins in serum
IgG 11.0 |
respectiveof% concentrationserum adultin |
|
|
IgA |
2.25 |
100 |
|
IgM |
1.15 |
|
|
IgD |
0.03 |
50 |
|
IgE |
0.0002 |
|
|
|
|
|
|
|
g/L |
–3 |
|
|
|
|
|
|
From Mother |
|
|
IgM |
|
|
|
|
|
|
|
IgG |
|
|
|
|
|
|
IgA |
|
|
|
|
|
|
IgD |
|
|
|
|
|
|
IgE |
|
3 |
6 |
9 |
12 |
15 |
18 |
93 |
Birth |
|
Age (months) |
|
(After Hobbs) |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

4 Blood
94
Immune System
Fundamental Principles
The body has nonspecific (innate) immune defenses linked with specific (acquired) immune defenses that counteract bacteria, viruses, fungi, parasites and foreign (non-self) macromolecules. They all function as antigens, i.e., substances that stimulate the specific immune system, resulting in the activation of an- tigen-specific T lymphocytes (T cells) and B lymphocytes (B cells). In the process, B lymphocytes differentiate into plasma cells that secrete antigen-specific antibodies (immunoglobulins, Ig) (!C). Ig function to neutralize and opsonize antigens and to activate the complement system (!p. 96). These mechanisms ensure that the respective antigen is specifically recognized, then eliminated by relatively nonspecific means. Some of the T and B cells have an immunologic memory.
Precursor lymphocytes without an antigenbinding receptor are preprocessed within the thymus (T) or bone marrow (B). These organs produce up to 108 monospecific T or B cells, each of which is directed against a specific antigen. Naive T and B cells which have not previously encountered antigen circulate through the body (blood !peripheral lymphatic tissue !lymph !blood) and undergo clonal expansion and selection after contact with its specific antigen (usually in lymphatic tissue). The lymphocyte then begins to divide rapidly, producing numerous monospecific daughter cells. The progeny differentiates into plasma cells or “armed” T cells that initiate the elimination of the antigen.
Clonal deletion is a mechanism for eliminating lymphocytes with receptors directed against autologous (self) tissue. After first contact with their specific self-antigen, these lymphocytes are eliminated during the early stages of development in the thymus or bone marrow. Clonal deletion results in central immunologic tolerance. The ability of the immune system to distinguish between endogenous and foreign antigens is called self/nonself recognition. This occurs around the time of birth. All substances encountered by that time are recognized as endogenous (self); others are identified as foreign (nonself). The inability to distinguish self from nonself results in autoimmune disease.
At first contact with a virus (e.g., measles virus), the nonspecific immune system usually cannot prevent the viral proliferation and the development of the measles. The specific immune system, with its T-killer cells (!B2) and Ig (first IgM, then IgG; !C3), responds slowly: primary immune response or sensitization. Once activated, it can eliminate the pathogen, i.e., the individual recovers from the measles.
Secondary immune response: When infected a second time, specific IgG is produced much more rapidly. The virus is quickly eliminated, and the disease does not develop a second time. This type of protection against infectious disease is called immunity. It can be achieved by vaccinating the individual with a specific antigen (active immunization). Passive immunization can be achieved by administering ready-made Ig (immune serum).
Nonspecific Immunity
Lysozyme and complement factors dissolved in plasma (!A1) as well as natural killer cells (NK cells) and phagocytes, especially neutrophils and macrophages that arise from monocytes that migrate into the tissues (!A2) play an important role in nonspecific immunity. Neutrophils, monocytes, and eosinophils circulate throughout the body. They have chemokine receptors (e.g., CXCR1 and 2 for IL- 8) and are attracted by various chemokines (e.g., IL-8) to the sites where microorganisms have invaded (chemotaxis). These cells are able to migrate. With the aid of selectins, they dock onto the endothelium (margination), penetrate the endothelium (diapedesis), and engulf and damage the microorganism with the aid of lysozyme, oxidants such as H2O2, oxygen radicals (O2–, OH·, 1O2), and nitric oxide (NO). This is followed by digestion (lysis) of the microorganism with the aid of lysosomal enzymes. If the antigen (parasitic worm, etc.) is too large for digestion, other substances involved in nonspecific immunity (e.g., proteases and cytotoxic proteins) are also exocytosed by these cells.
Reducing enzymes such as catalase and superoxide dismutase usually keep the oxidant concentration low. This is often discontinued, especially when macrophages are activated (!below and B3), to fully exploit the bactericidal effect of the oxidants. However,
Despopoulos, Color Atlas of Physiology © 2003 Thieme |
! |
All rights reserved. Usage subject to terms and conditions of license. |
|

A. Nonspecific immune defenses enhanced by specific antibodies |
|
|
|
|
|
|
|
|||||||
|
|
|
Humoral |
Cellular |
|
|
|
|
|
|
|
|
|
|
Lysozyme |
|
|
|
|
|
|
Fc and C3b |
|
|
|
||||
|
|
|
|
|
|
receptors |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|||
Damages membranes |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Interferons (IFN) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IFN-α, β, γ inhibit |
|
|
|
|
Neutrophils, |
|
|
|
|
|
|
|
|
|
viral proliferation; |
|
|
|
|
Monocytes |
|
|
n |
|
|
|
Ig |
|
|
|
|
|
|
|
2 |
ige |
|
|
|
|
|
|||
IFN-γ activates macrophages, |
|
|
|
→ Macrophages |
|
Ant |
|
C3b |
|
|
||||
|
killer cells, |
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
System |
|
|
B and T cells |
|
|
|
|
|
Antigens opsonized |
|
||||||
Complement activation |
|
|
|
Phagocytosis |
by Ig and C3b |
|
||||||||
|
|
|
|
|
|
|
|
|
|
|||||
|
|
1 |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Alternative |
Classical |
|
|
|
|
|
|
|
|
|
|
|
Immune |
|
|
|
|
|
Lysis |
|
|
|
|
|
|
||||
C3 |
C1q |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
Release of: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidants, |
|
|
Membrane |
|
|||||
|
Antigen-antibody |
|
|
|
|
|
||||||||
|
complexes |
|
|
|
proteases, |
|
|
damage |
|
4.4 |
||||
|
|
|
|
mediators of |
|
|
|
|
|
|
|
|
||
Micro- |
|
|
|
|
|
|
Activated |
|
||||||
C1q |
|
|
|
inflammation |
|
|
|
Plate |
||||||
organisms |
|
|
|
|
|
|
||||||||
|
|
n |
|
|
|
|
macrophages |
|
||||||
|
|
|
|
|
|
|
(see plate B3) |
|
||||||
|
|
ige |
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
t |
|
|
|
|
|
|
|
|
|
|
|
||
|
n |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
A |
|
|
C3a |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C4a |
|
Inflammation |
|
|
|
|
|
|||
|
|
|
|
C5a |
|
|
|
|
|
|
||||
C3b |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Opsonization |
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
||||
|
of antigen |
|
|
|
|
Mediators of |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|||||
|
A |
|
|
|
Activation |
|
|
inflammation |
|
|
||||
|
nti |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
g |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
e |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
n |
|
|
|
|
|
|
|
|
|
|
|
Membrane attack |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
complex (C5–C9) |
|
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
Mast cells, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
basophils |
|
|
|
|
|
|
n |
|
|
|
Na+ |
|
|
|
|
|
|
|
ntig |
e |
|
|
||
|
|
3 |
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
A |
|
|
|
|
||||
|
H2O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidants |
|
|
Eosinophils |
|
|
|
|
|
|
|
|
|
|
|
Proteases |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
Na+ |
|
Perforins |
|
|
|
|
|
|
|
|
|
|
|
Antigen: |
H2O |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
Natural |
|
|
|
|
|
|
|
|
|
|
|
Pathogen, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
foreign cell, |
|
|
|
killer cell |
|
|
|
|
|
|
|
|
|
|
virus-infected |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
endogenous cell |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ADCC |
|
|
|
|
|
|
|
|
|
|
IgA |
|
||
Cytolysis |
|
|
|
|
|
|
IgE |
IgE IgM |
IgG |
|
||||
|
Fc receptor |
|
|
IgG |
|
|
|
|
|
|
|
|||
|
|
|
|
|
Fc |
|
|
|
|
|
|
|
|
95 |
|
|
|
|
|
IgG |
|
|
Immunglobulins |
||||||
|
|
|
|
|
|
|
(see plate C3) |
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|||||
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

!
the resulting inflammation (!A2, 4) also damages cells involved in nonspecific defense and, in some cases, even other endogenous cells.
|
Opsonization (!A1, 2) involves the binding of |
|
|
opsonins, e.g., IgG or complement factor C3b, to |
|
|
specific domains of an antigen, thereby en- |
|
|
hancing phagocytosis. It is the only way to |
|
|
make bacteria with a polysaccharide capsule |
|
|
phagocytable. The phagocytes have receptors |
|
|
on their surface for the (antigen-independent) |
|
|
Fc segment of IgG as well as for C3b. Thus, the |
|
|
antigen-bound IgG and C3b bind to their re- |
|
|
spective receptors, thereby linking the rather |
|
|
unspecific process of phagocytosis with the |
|
|
specific immune defense system. Carbohy- |
|
|
drate-binding proteins (lectins) of plasma, |
|
|
called collectins (e.g. mannose-binding pro- |
|
|
tein), which dock onto microbial cell walls, also |
|
Blood |
acts as unspecific opsonins. |
|
The complement cascade is activated by an- |
||
tigens opsonized by Ig (classical pathway) as |
||
|
||
4 |
well as by non-opsonophilic antigens (alterna- |
|
tive pathway) (!A1). Complement com- |
||
|
||
|
ponents C3a, C4a and C5a activate basophils |
|
|
and eosinophils (!A4). Complement com- |
|
|
ponents C5 –C9 generate the membrane-attack |
|
|
complex (MAC), which perforates and kills |
|
|
(Gram-negative) bacteria by cytolysis (!A3). |
|
|
This form of defense is assisted by lysozyme |
|
|
(= muramidase), an enzyme that breaks down |
|
|
murein-containing bacterial cell walls. It oc- |
|
|
curs in granulocytes, plasma, lymph, and |
|
|
secretions. |
|
|
Natural killer (NK) cells are large, granular |
|
|
lymphocytes specialized in nonspecific |
|
|
defense against viruses, mycobacteria, tumor |
|
|
cells etc. They recognize infected cells and |
|
|
tumor cells on “foreign surfaces” and dock via |
|
|
their Fc receptors on IgG-opsonized surface |
|
|
antigens (antibody-dependent cell-mediated |
|
|
cytotoxicity, ADCC; !A3). Perforins exocytosed |
|
|
by NK cells form pores in target cell walls, |
|
|
thereby allowing their subsequent lysis (cy- |
|
|
tolysis). This not only makes the virus unable to |
|
|
proliferate (enzyme apparatus of the cell), but |
|
|
also makes it (and other intracellular patho- |
|
|
gens) subject to attack from other defense |
|
|
mechanisms. |
|
96 |
Various interferons (IFNs) stimulate NK cell activity: |
|
IFN-α, IFN-" and, to a lesser degree, IFN-γ. IFN-α and |
||
|
IFN-" are released mainly from leukocytes and fibro- |
blasts, while IFN-γ is liberated from activated T cells and NK cells. Virus-infected cells release large quantities of IFNs, resulting in heightened viral resistance in non-virus-infected cells. Defensins are cytotoxic peptides released by phagocytes. They can exert unspecific cytotoxic effects on pathogens resistant to NK cells (e.g., by forming ion channels in the target cell membrane).
Macrophages arise from monocytes that migrate into the tissues. Some macrophages are freely mobile (free macrophages), whereas others (fixed macrophages) remain restricted to a certain area, such as the hepatic sinus (Kupffer cells), the pulmonary alveoli, the intestinal serosa, the splenic sinus, the lymph nodes, the skin (Langerhans cells), the synovia (synovial A cells), the brain (microglia), or the endothelium (e.g., in the renal glomeruli). The mononuclear phagocytic system (MPS) is the collective term for the circulating monocytes in the blood and macrophages in the tissues. Macrophages recognize relatively unspecific carbohydrate components on the surface of bacteria and ingest them by phagocytosis. The macrophages have to be activated if the pathogens survive within the phagosomes (!below and B3).
Specific Immunity: Cell-Mediated Immune Responses
Since specific cell-mediated immune responses through “armed” T effector cells need a few days to become effective, this is called delayed-type immune response. It requires the participation of professional antigen-pres- enting cells (APCs): dendritic cells, macrophages and B cells. APCs process and present antigenic peptides to the T cells in association with MHC-I or MHC-II proteins, thereby delivering the co-stimulatory signal required for activation of naive T cells. (The gene loci for these proteins are the class I (MHC-I) and class II (MHC-II) major histocompatibility complexes (MHC)), HLA (human leukocyte antigen) is the term for MHC proteins in humans. Virusinfected dendritic cells, which are mainly located in lymphatic tissue, most commonly serve as APCs. Such HLA-restricted antigen presentation (!B1) involves the insertion of an antigen in the binding pocket of an HLA protein. An ICAM (intercellular adhesion molecule) on the surface of the APC then binds to
!
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.

B. Specific immunity: T-cell activation |
|
|
|
|
|
|||
|
|
|
|
|
T lymphocytes |
|
|
|
Antigen-presenting cells: |
CD8+ T cells recognize |
CD4+ T cells recognize |
|
|||||
antigen in HLA(MHC)-I |
antigen in HLA(MHC)-II |
|
||||||
– Macrophages |
|
|
||||||
|
|
|
|
|
|
|
||
– Dendritic cells |
|
|
|
|
|
|
|
|
– B cells |
|
|
|
|
|
|
|
|
|
|
APC |
|
Antigen |
|
T cell |
|
|
|
|
HLA-I |
|
CD8 or CD4 |
“Naive” |
II |
||
|
|
|
System |
|||||
|
|
or HLA-II |
|
T-cell receptor |
T cell |
|||
|
|
|
|
|
|
Dual signal |
|
|
|
|
|
B7 |
|
CD28 |
|
|
|
|
|
|
|
|
|
Immune |
||
Example: |
1 |
|
|
|
|
|
||
Dendritic cell |
ICAM |
|
LFA1 |
|
|
|||
|
|
|
|
|||||
|
|
|
|
|
IL-2 |
|
|
|
|
|
|
|
|
|
|
4.5 |
|
|
|
|
|
|
IL-2 receptor |
|
||
|
|
|
|
|
|
|
|
Plate |
|
|
|
|
T-cell proliferation |
|
|
||
|
|
|
(clonal expansion and differentiation) |
|
|
|||
|
|
|
CD8/HLA-I |
CD4/HLA-II |
|
|
||
Infected cell, |
|
|
|
|
|
TH2 cell |
|
|
tumor cell, foreign cell |
|
Cytotoxic T cell |
TH1 cell |
|
|
|||
|
|
|
IL-10, |
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IFNγ |
TGFβ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
See plate C |
|
Infected |
|
|
LFA1 |
T cell |
|
|
|
|
cell |
ICAM |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
HLAI |
|
CD8 |
|
|
|
|
|
|
|
T-cell receptor |
Macrophage |
|
|
|||
|
|
|
|
|
||||
Antigen |
|
|
|
|
activation |
|
|
|
|
|
Fas ligand |
|
|
|
|
|
|
|
CD95 |
|
|
3 |
|
|
|
|
Apoptosis |
|
Perforins |
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
Granzyme B |
|
Macrophage |
|
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Inflammation |
|
97 |
|
|
|
|
|
|
|
|
||
Despopoulos, Color Atlas of Physiology © 2003 Thieme
All rights reserved. Usage subject to terms and conditions of license.