

теза
в иле, взятом в русле Волги, с
повышением
температуры от 0,5 до
20°С
увеличивалась с 0,95 до
3,09
мг/л С в -сутки, в иле из поймы —
с
1,61 до 5,65 мг/л С в сутки (Соро-
кин,
1958).
Интенсивность
хемосинтеза уста-
навливают
путем количественного
учета
специфических продуктов, об-
разующихся
в результате работы ,ав-
тотрофных
бактерий или определени-
ем
количества СОг, идущего на син-
тез
органического вещества. В послед-
нем
случае используют, радиоуглерод-
ный
метод.
Недавно
выявлена огромная роль
хемосинтезирующих
бактерий в окис-
лении
ювенильного сероводорода в
глубоководных
гидротермах рифтовых
зон
(см. гл. 2). Численность таких
бактерий
близ выхода термальных
вод
может достигать 5 млн./мл (Каг1
е!
а11., 1980).
Энергобаланс
экосистем. В любую
экосистему
непрерывно поступает энергия —
солнечная, химиче-
ская,
механическая или иная. В ходе экосистемных
процессов она
связывается,
трансформируется или остается интактной
(напри-
мер,
энергия многих пластмасс и других
стойких химических со-
единений).
Использованная гидробионтами энергия
рассеивается
в
экосистеме, аккумулируется в ней
(прирост биомассы, накопле-
ние
энергоемких соединений) или мигрирует
с биогеопотоками
(выход
организмов из воды, их вылов, выделение
энергоемких га-
зов,
вынос энергии со стоком). Во всех случаях
в соответствии с
законами
термодинамики количество энергии,
поступившей на
вход,
равно алгебраической сумме ее приращения
в системе и вы-
хода
из системы.
Как
показал Е. Шредингер (1944), для поддержания
внутрен-
ней
упорядоченности в системе необходима
постоянная работа по
«выкачиванию»
неупорядоченности. В экосистеме
отношение
дыхания
к суммарной биомассе можно рассматривать
как отно-
шение
затрат энергии на поддержание
жизнедеятельности к энер-
гии,
заключенной в структуре, или как меру
термодинамической
упорядоченности.
Это соотношение Шредингера
одновременно
служит
и мерой скорости экологического оборота
(отношение
«пропускания»
к «содержанию»). Если выразить дыхание
и био-
массу
в энергетических величинах и разделить
их на абсолютные
температуры,
то рассматриваемое отношение становится
отноше-
нием
прироста энтропии (и соответствующей
работы), связанного
с
обеспечением структуры, к энтропии
упорядоченной части.
О
50 100
Рис.
60. Относительная ве-
личина
хемосинтеза на раз-
ных
глубинах Рыбинского
водохранилища
в процентах
от
наблюдающейся в при-
донном
слое (по Сорокину,
1958)
354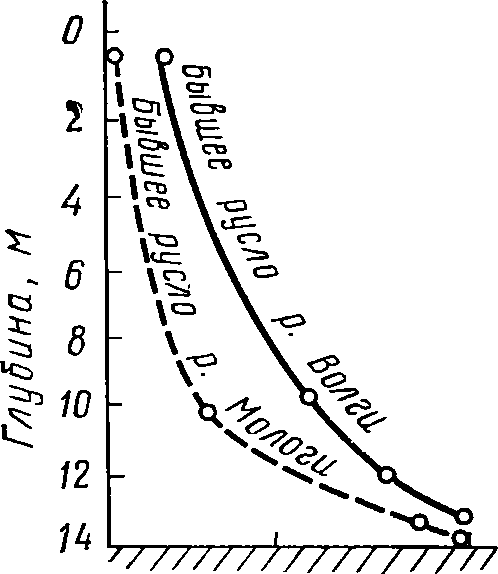
Относительная Величина хемосинтеза, %
Рис.
61. Трансформация веществ и энергии в
гидробиоценозах (по Одуму, 1968,
и
Крохину, 1969). А
— морской залив Лонг-Айленд (США); Б
— оз. Дальнее
(Камчатка):
а
—
фитопланктон, б
—
бактерии, в
— фкльтраторы,
г
— хищный
зоопланктон, д
— колюшка,
е
— молодь
нерки, ж
— голец
Цифры
в заштрихованных фигурах — биомасса
организмов в кДж/м2,
цифры в
каналах
— поток энергии в кДж/м2-год
Наибольшее
значение в балансе экосистем имеет
связывание
солнечной
энергии, поступление и вынос энергоемких
веществ,
изменение
биомассы и количества энергоемких
веществ в системе.
Меньшее
значение имеет поступление тепловой
и механической
энергии
(приливы и отливы, ветровые воздействия,
Теплообмен
вод
и др.). В связи с этим, а также по причине
трудности учета
миграция
тепловой и механической энергии в
энергобалансе эко-
систем
не рассматривается.
В
экосистемах разного типа энергобаланс
зависит от особен-
ностей
их живого и косного компонентов. На
рис. 61 показана
схема
энергопотока в заливе Лонг-Айленд и
оз. Дальнем (Камчат-
ка).
Из всей энергии, связываемой
фотосинтетиками залива, 50%
расходуется
на дыхание, 50% аккумулируется в
энергоемких сое-
динениях.
Организмы зоопланктона рассеивают
25% аккумулиро-
ванной
энергии, обитатели дна—15, вторичные
консументы —
6%.
В Северном море в столбе воды с основанием
1 м2
в расте-
ниях
в среднем за год образуется 100 г углерода,
а вылов рыбы
достигает
0,22 г С/м2
(51ее1е, 1965). Как правило, с переходом
на
следующий трофический уровень
рассеивается 80—90% исход-
ной
энергии. Наибольшее количество ее
энтропизируется расти-
тельноядными
животными (30—40%), продуцентами (20—30%)
и
редуцентами (20—40%). Особенно велико
относительное рас-
сеивание
энергии бактериями, биомасса которых
в биоценозе
составляет
лишь небольшую часть общей, а энерготраты
— 20—
40%
и более. Например, в оз. Марион (Канада)
из общего коли-
12*
355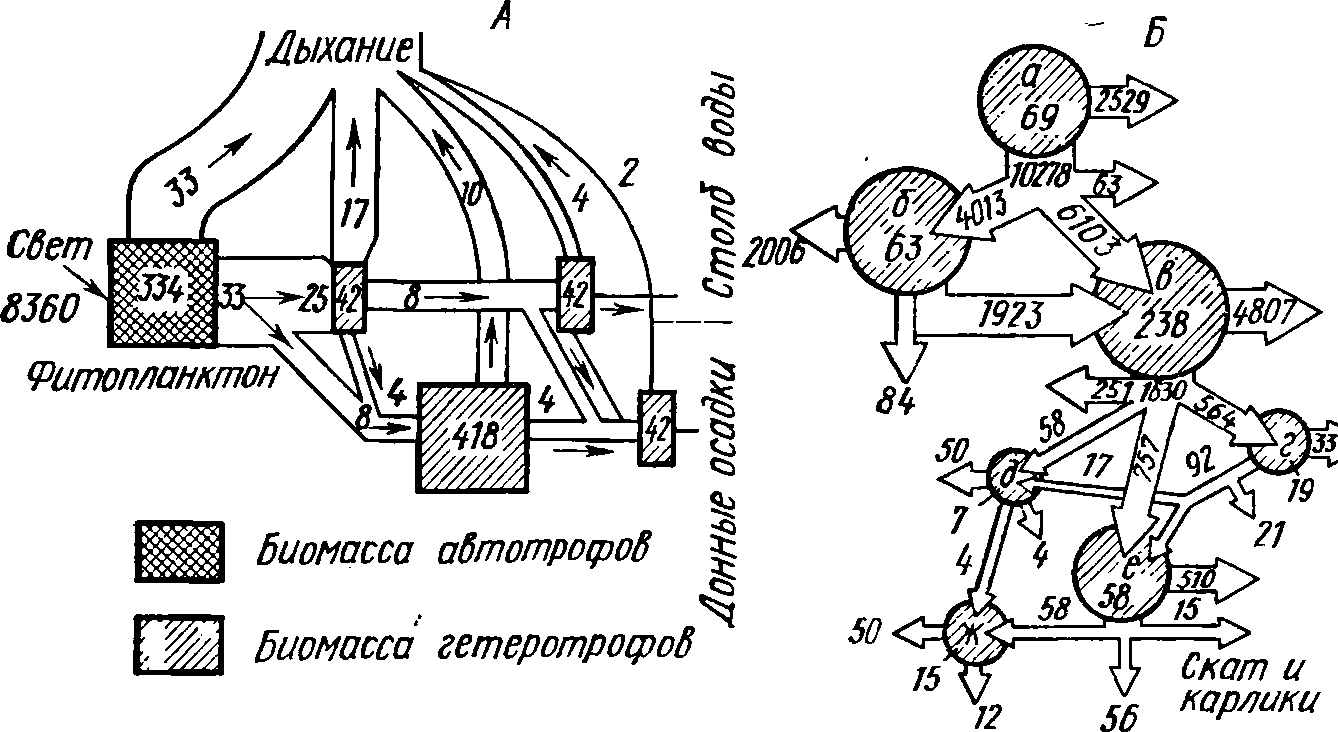
чества
потребленного донными организмами
кислорода на долю
бактерий
приходилось летом 30%, зимой 45%.
В
оз. Байкал положительная часть его
энергобаланса сла-
гается
(в кт С в год) из первичной продукции
(3 951), аллохтон-
ной
органики, приносимой притоками (292) и
осадками (12).
Расход
связан с выносом органики Ангарой
(74), деструкцией
(4
054), захоронением в донных осадках
(126), выловом рыбы и
нерпы
(1). Фитопланктон утилизирует за год
0,11% падающей
солнечной
радиации (Вотинцев и др., 1975).
В
Рыбинском водохранилище на первом
трофическом уровне
аккумулируется
за год на 1 м2
в процессе фотосинтеза 2213 и
1344
кДж в виде бактерий, развивающихся за
счет органических
веществ,
поступающих в водохранилище извне. В
бактерии, зоо-
планктон
и зообентос второго трофического
уровня переходят
соответственно
441, 328, 55 кДж. В процессе дыхания
перечислен-
ными
группами организмов рассеиваются (в
том же порядке)
1323,
286, 10 кДж или всего 1710 кДж. В органическом
веществе
консументов
второго порядка — хищных планктофагов
и бентофа-
гов,
личинок рыб и мирных рыб аккумулируется
соответственно
15,
17, 25, 50 кДж, а всего 152 кДж. Траты на
дыхание перечислен-
ных
консументов составляли (в том же
порядке) 2, 5, 1, 7, 80, а
всего
106,7 кДж. На четвертом трофическом уровне
накапливается
соответственно
8,4; 4,2 и 12,6 кДж, рассеивается 10,1; 2,5 и
40,3
кДж — всего 52,9 кДж. Хищные рыбы, стоящие
на пятом
трофическом
уровне, аккумулируют 2,6 кДж, рассеивают
6,3 кДж.
Энергия,
накапливаемая организмами 1, 2, 3, 4 и
5-го трофических
уровней,
выражалась отношением 100:22,3:0,7:0,06.
Консументами
различных
порядков аккумулировалось от 9 до 23%
органических
веществ,
синтезируемых организмами предыдущих
трофических
уровней.
За вегетационный период 1971 г. продукция
фи-
топланктона
равнялась 55 г С/м2
( что соответствовало использо-
ванию
0,12% общей проникающей солнечной
энергии), продукция
бактерий
— 30 г. Деструкция органического вещества
в толще
воды
достигала 79 г С, а за счет аэробных и
анаэробных процес-
сов
в иловых отложениях — соответственно
20 и 48 гС/м2
(С.
И. Кузнецов и др., 1974).
Динамика
экосистем
Ни
одна экосистема не существует вечно
и рано или поздно
сменяется
другой, причем в этом процессе
обнаруживается опре-
деленная
необратимая направленность. Такие
закономерные с за-
ранее
предсказуемым результатом изменения
называются сукцес-
сиями.
Различают первичные
и вторичные
сукцессии.
Первые ха-
рактеризуют
становление и развитие экосистем во
вновь обра-
зуемых
водоемах, вторые — постепенное изменение
длительно
существующих
экосистем.
В
процессе сукцессии биоценоз приближается
к состоянию,
наиболее
соответствующему абиотической среде,
к состоянию
наибольшей
устойчивости экосистемы — ее климаксу.
Оно может
356
нарушаться
переменами в абиотической среде, и
тогда биоценоз
снова
претерпевает последовательные
изменения, отражающие
саморегуляторные
свойства системы. Весь градиент
сообществ,
сменяющих
друг друга до достижения климакса,
называется се-
рией.
Сериальные
изменения сопровождаются стабилизацией
са-
мой
системы в смысле достижения максимальной
защиты от
возмущающих
воздействий среды. Сукцессия может
происходить
вследствие
изменения биотопа самим биоценозом
(автогенная
сукцессия)
или внешними силами (аллогенная,
или экзогенная,
сукцессия),
например в результате деятельности
человека. В от-
личие
от аллогенной автогенная сукцессия
всегда идет в направ-
лении
эволюционного развития биосферы —
повышения ее гомео-
стаза,
увеличения со временем контроля над
физическими фак-
торами
среды и растущей независимости от них.
Помимо
сукцессий, для экосистем характерны
закономерные
изменения
состояния, сопровождающиеся направленной
перестрой-
кой,
но имеющие обратимый характер,—
флуктуации.
В этом слу-
чае
структурно-функциональная организация
экосистемы, видоиз-
меняясь,
в частности, на протяжении года, в общем
имеет один
и
тот же облик в каждый сезон на протяжении
многих лет.
Сукцессия
как экосистемный процесс. Гидробиоценоз
как сба-
лансированная
система может существовать только при
условии,
когда
продукты метаболизма входящих в него
популяций не нака-
пливаются
в биотопе и не изменяют его характеристик,
в соответ-
ствии
с которыми находятся структура и
функциональные особен-
ности
сообщества. Если продукты метаболизма
накапливаются в
биотопе,
происходит его «загрязнение», т. е.
ухудшение условий
существования
для обитающих здесь гидробионтов.
Вместе с тем
биотоп
становится более благоприятным
местообитанием для дру-
гих
организмов. В результате изменяется
структура и, соответст-
венно,
функциональные особенности биоценоза.
Он будет стабиль-
нее,
если жизнедеятельность входящих в
него популяций будет
организована
по типу «безотходной технологии». Чем
быстрее
биоценоз
изменяет свой биотоп — условия своего
существования,
тем
скорее меняется сам.
Стабильность
биоценоза возможна и при
несбалансированно-
сти
его обмена, не сопровождающейся
изменением биотопа и на-
коплением
в нем продуктов метаболизма, меняющих
условия
существования
популяций. Так, достаточно быстрое
течение исклю-
чает
накопление метаболитов в реках. Еще
слабее влияние биоце-
нозов
на водную толщу Мирового океана. Внешние
воздействия
на
экосистему могут ускорять, замедлять
или исключать ее сук-
цессию
в зависимости от того, насколько
вызываемые ими изме-
нения
биотопа совпадают с теми, которые
являются результатом
биоценотических
процессов. Например, биотическое
заиление дна
может
ослабляться промыванием водоема или
усиливаться за
счет
оседания частиц, приносимых извне.
Среди
водоемов наиболее выраженной сукцессией
характери-
зуются
озера, часто превращающиеся из
олиготрофных в мезо-
357
о
о
Рис.
62. Изменение экосистем озер по данным
палеонтологии (по Оаи1с1еп, 1966).
А,
Б
— изменение видового разнообразии
(индекс Шеннона — Н)
сообществ хи-
дорид
в оз. Монтерози, В
— изменение соотношения количества
босмин и хидо-
рид
(в %) в оз. Лагуна-де Петенксиль
и
эвтрофные. Вследствие непрерывного
накопления биогенных
осадков,
усиливаемого осаждением взвешенного
материала, при-
носимого
реками, озеро постепенно мелеет, объем
гиполимниона
уменьшается,
кислородный режим ухудшается, количество
биоге-
нов
возрастает; население озера становится
количественно богаче,
так
как вследствие повышения концентрации
биогенов первопищи
образуется
больше. Одновременно из фауны и флоры
выпадают
представители,
наиболее требовательные к чистоте
воды. Биоген-
ное
илонакопление и количественное
обогащение населения начи-
нают
ускоряться, сдвиги в начавшемся
направлении усиливаются,
озеро
мелеет еще больше, подводные растения
прибрежной полосы
(рдесты,
уруть, роголистник и др.) получают
возможность рас-
пространяться
все дальше к центру, пока не покроют
всю площадь
дна.
Жизнь количественно становится еще
богаче, еще быстрее идет
накопление
донных отложений за счет отмирающих
растений и
животных.
Начинается наступление надводных
растений (тростник,
камыш,
рогоз и др.) и постепенное заболачивание
водоема, завер-
шающееся
его исчезновением.
Реальная
картина сукцессии озер очень сильно
отличается от
приведенной
схемы и зависит от конкретных проявлений
во вре-
мени
внешних сил, воздействующих на экосистему
(изменение
климатических,
гидрологических и других факторов).
Сукцессия
может
задерживаться на каких-то стадиях,
прерываться или даже
в
отдельных случаях протекает в обратном
направлении, когда
результаты
биологических процессов перекрываются
абиотиче-
скими
или антропогенными воздействиями.
Однако само развитие
экосистем,
соответствующее закономерностям их
существования,
есть
определенным образом направленный
процесс, подчиняю-
щийся
общим положениям термодинамики и
законам преобразо-
вания
систем.
358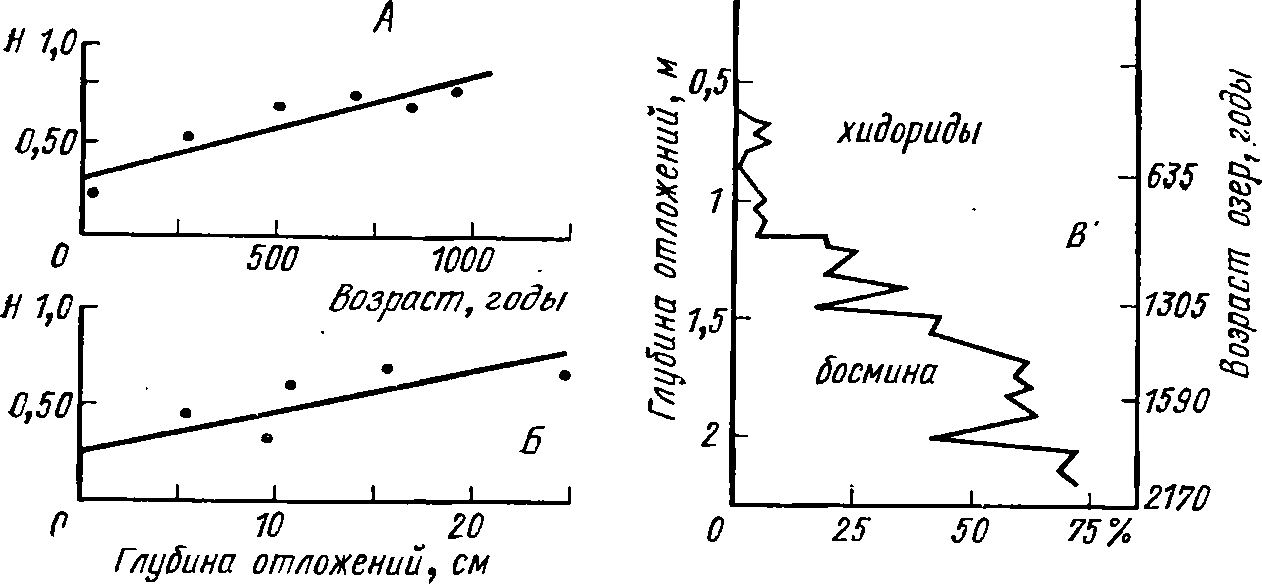
Один
из надежных способов выявления истинной
картины дли-
тельного
изменения озерной экосистемы — анализ
остатков жи-
вотных
и растений из разных горизонтов донных
отложений, воз-
раст
которых может быть определен с
достаточной точностью.
Устанавливая
по сохранившимся фрагментам видовой
состав и
численность
популяций отдельных групп организмов
(диатомовые
водоросли,
ракообразные, личинки насекомых и
др.), можно ре-
конструировать
облик экосистем на тех или иных временных
раз-
резах
и судить о его эволюции. Например, при
изучении оз. Па-
лецкого
(Московская обл.) установлено, что после
накопления
на
его дне метровой толщи осадков произошла
резкая смена
структуры
сообщества. В частности, преобладание
рачков А1опа
ацШз
и А1опе11а
папа
сменилось доминированием Скуйогиз
зркаегьсиз
и А1опейа
ехыза
(Смирнов, 1971). На рис. 62 видно,
как
в течение двух тысяч лет в оз.
Лагуна-де-Петенксиль (Цен-
тральная
Америка) снижается количество босмин
(глубоковод-
ные
формы) и возрастает роль хидорид
(обитатели мелководья,
зароелевые
формы). На том же рисунке хорошо
прослеживается
усложнение
экосистемы (рост видового разнообразия)
на протя-
жении
1000 лет существования оз. Монтерози
(Италия). Исполь-
зуя
палеолимнологический метод (диатомовый,
карцинологиче-
ский
и другие анализы), следует учитывать
границы его воз-
можностей.
В субфосильных отложениях многие
фрагменты часто
не
сохраняются, не все находимые остатки
организмов опреде-
ляются
до вида, возможно нарушение стратиграфии
вследствие
взмучивания
грунта течениями и его перекапывания
животными.
Кульминацией
развития экосистем (климаксом) считается
со-
стояние,
при котором биомасса на единицу потока
энергии дости-
гает
максимума. Отношение затрат энергии
на поддержание жиз-
недеятельности
к энергии, заключенной в структуре,
снижается,
т.
е. уменьшается отношение энергии
дыхания сообщества к той,
которая
связана в его биомассе. Такой результат,
в частности,
достигается
укрупнением организмов, сопровождающимся
сниже-
нием
интенсивности дыхания. Отношение
валового фотосинтеза к
■суммарному
дыханию сообщества стремится к 1, т. е.
приход и
расход
энергии уравновешивают друг друга.
Следовательно, выход
чистой
продукции (превышение фотосинтеза над
дыханием) приб-
лижается
к 0, к 0 стремится и выход продукции на
единицу био-
массы.
В
процессе сукцессии структурные
изменения экосистемы про-
являются
прежде всего в усложнении организации
биоценоза.
Возрастает
число видов, повышается их эквитабильность,
которая
довольно
скоро стабилизируется, а на поздних
стадиях даже сни-
жается.
Усиливается стратификация, в результате
чего в сообще-
стве
появляются новые экологические ниши.
Непрерывно возра-
стает
разнообразие веществ, выделяемых
компонентами биоценоза
в
воду, как побочных продуктов возросшего
метаболизма сооб-
щества.
В связи с этим возрастает роль метаболитов
как регуля-
тора,
стабилизирующего состояние экосистемы.
Общее количество
359
органического
вещества в системе повышается, пищевые
цепи из
линейных,
в основном пастбищных с преобладанием
фитофагов,
становятся
разветвленными, преимущественно
детритными. Жиз-
ненные
циклы организмов удлиняются и
усложняются, возрастает
специализация
по нишам. Иногда вследствие усиления
конкурен-
ции
и некоторых других причин видовое
разнообразие, сначала
возрастающее,
несколько снижается.
Для
пелагических сообществ в процессе
сукцессии описано
изменение
размерной структуры. На ее начальных
стадиях преоб-
ладают
мелкие, быстро размножающиеся организмы,
в дальней-
шем
повышается роль более крупных форм,
темп роста и скорость
размножения
которых ниже.
Основные
закономерности сукцессии экосистем
отражают об-
щий
ход их эволюции как блоков биосферы.
Каждый из множе-
ства
видов, входящих в биоценоз, по-своему
реагирует на измене-
ние
среды, но во избежание биогеоценотического
хаоса их эволю-
ция
должна быть согласованной, быть
синэволюцией. При этом
виды-доминанты,
относительно самостоятельные в
биоценозе, оп-
ределяют
темпы и формы эволюции видов-сателлитов.
Синэволю-
ция
видов наиболее отчетливо проявляется
в сходстве реакции
на
изменение среды, что обеспечивает
гармоничное развитие
сообщества
как целого при колебании внешних
условий. Поэтому
свойства
биоценоза отражаются в свойствах
образующих-его по-
пуляций,
особенно доминантных видов.
Поддержание
биогеоценотического равновесия
представляет
собой
результат совместной эволюционно
согласовайной работы
популяционных
и биоценотических механизмов.
Биологический
прогресс,
основанный преимущественно на повышении
морфо-
физиологической
организации, имеет следствием резкое
повыше-
ние
скорости трансформации веществ и
энергии. Биологический
прогресс,
основанный на совершенствовании
популяционной струк-
туры,
увеличивает организационное единство
биоценоза, повышает
уровень
его целостности. Единством обеих форм
прогресса яви-
лось
совершенствование биогеоценозов как
систем.
Своеобразна
тенденция к увеличению размера
гидробионтов
в
процессе сукцессии. Мелкие размеры
дают преимущества в
среде,
богатой растворенными веществами,
нужными организму
(увеличение
относительной поверхности). По мере
развития эко-
системы
и снижения концентрации биогенных
веществ преимуще-
ство
может переходить к более крупным
организмам с дли-
тельным
жизненным циклом, которые лучше
адаптированы к исг
пользованию
периодических (сезонных и др.) обогащений
среды
биогенами.
В энергетическом плане укрупнение
организмов, сопро-
вождающееся
снижением интенсивности дыхания,
создает возмож-
ность
существования за счет единицы поступающей
энергии все
большей
биомассы сообщества. Таким образом
реализуется одна
из
основных закономеоностей саморазвития
экосистем — снижение
отношения
пропускаемой энергии к энергии,
заключенной в био-
ценозе.
Живое вещество, экономнее используя
доступную энергию,
360
получает
возможность увеличивать свое количество
в биосфере.
На
эту закономерность в свое время обратил
внимание Ч. Дар-
вин,
говоря об «увеличении суммы жизни» в
процессе эволюции.
В
ходе сукцессии изменяются характер и
скорость круговорота
биогенных
веществ. На начальных стадиях развития
экосистем
круговорот
минеральных веществ открыт, роль
детрита в регене-
рации
биогенов незначительная, скорость
обмена между биоцено-
зом
и биотопом высокая. В дальнейшем
круговорот становится
более
закрытым, поскольку в значительной
степени замыкается
в
пределах биоценоза. Соответственно
этому скорость обмена меж-
ду
сообществом и биотопом снижается, роль
детрита, количество
которого
в экосистеме возрастает, увеличивается.
На зрелых ста-
диях
экосистемы более способны к захвату
и удержанию их в
круговороте.
С
развитием экосистемы повышается ее
гомеостаз, устойчивость
против
возмущающего действия внешних сил. С
одной стороны,
это
достигается кондиционированием среды,
смягчением стрессо-
вых
влияний ее на сообщество. С другой
стороны, само сообще-
ство
изменяется в направлении большей
устойчивости за счет
усложнения
и усиления биотических связей. При
этом отрицатель-
ные
взаимодействия (паразитизм, хищничество,
конкуренция и др.)
становятся
сбалансированнее, а положительные
(протокооперация,
мутуализм
и др.) складываются все чаще и их
функциональное
значение
в экосистеме возрастает.
В
тех случаях, когда в результате
сериальных изменений отно-
сительное
количество автотрофов в биоценозе
снижается, говорят
об
автотрофной
сукцессии,
в обратном случае — о гетеротрофной.
Сущность
автотрофной сукцессии хорошо выражена
лабора-
торной
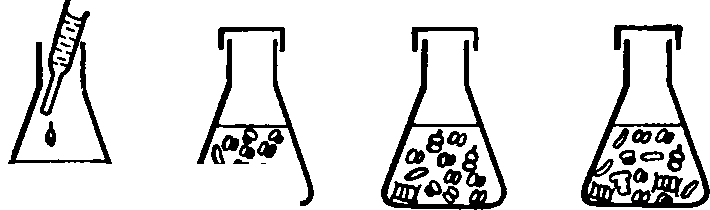
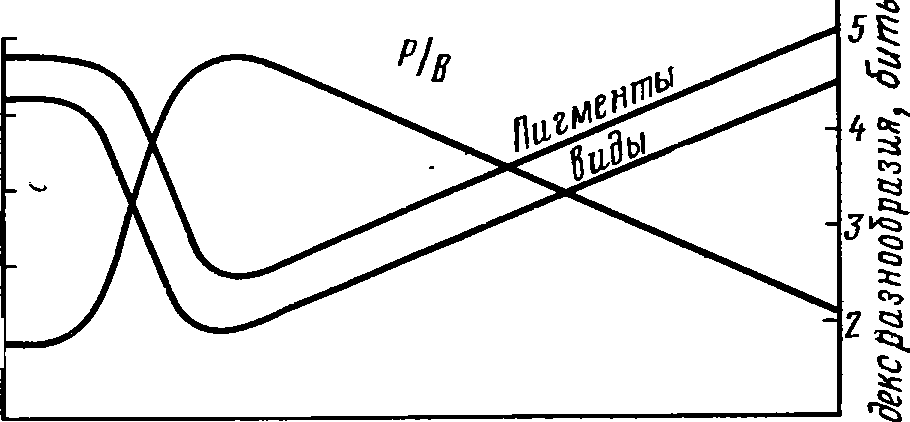
моделью, описанной Р. Марголефом (рис.
63). В колбе
со
старой культурой водорослей имеется
большое число видов
диатомовых,
зеленых, жгутиковых, перидиниевых,
коловраток.
Столь
же велико и биохимическое разнообразие
водорослей, о чем
говорит
богатство растительных пигментов,
обнаруживаемых в
колбе.
Количество энергии, связываемой
водорослями и рассеивае-
мой
обитателями колбы, почти одинаково,
прирост их, отнесенный
к
биомассе, мал. Если в колбу со старой
культурой добавить све-
жей
среды, картина резко меняется. В культуре
начинают преоб-
ладать
несколько видов зеленых водорослей,
следовательно, ин-
формация
биоценоза снижается. Точно так же
снижается разнооб-
разие
растительных пигментов и одновременно
резко увеличи-
вается
отношение прироста биомассы биоценоза
к ее исходной
величине.
Количество энергии, рассеиваемой в
биоценозе, стано-
вится
заметно меньше того, какое связывается
фотосинтетиками.
В
последующем все перечисленные показатели
постепенно ме-
няются,
приближаясь к тем, какими характеризовалась
старая
культура.
Через несколько недель сериальные
изменения закан-
чиваются—
достигается стадия климакса.
Моделью
гетеротрофной сукцессии может служить
колба с
сенным
настоем или иной органической средой.
Вначале в ней
361
Ш
(&к
Старая
Молодая
культура
* культура
Стареющие
культуры
^
га V)
^
Йс
Швг
411
4
Время
(около трех недель)
Рис.
63. Модель экологической сукцессия (по
Р. Марголефу).
В
колбах показаны изменения видового
Состава в течение сук-
цессии,
вызванной добавлением свежей1
питательной среды в
старую
«климаксную» культуру. Графир отражает
изменение
разнообразия
и отношения продукции к биомассе (по
Одуму,
1968):
/
— диатомовые, 2
—
зеленые водоросли, 3
— динофлагелляты, 4
— колеи
вратки
развиваются
гетеротрофные бактерии, затем появляются
простей-
шие,
в первую очередь инфузории, и другие
животные. После
значительного
снижения концентрации в воде органических
ве-
ществ
в культуре появляются водоросли. Их
число и разнообразие
благодаря
заносу извне всевозможных зачатков
постепенно воз-
растает,
процессы фотосинтеза усиливаются и,
наконец, начинают
уравновешивать
затраты энергии на дыхание. Достигается
состоя-
ние,
напоминающее климакс природных
гидробиоценозов.
Автотрофная
сукцессия. Такая форма сериальных
изменений
характерна
для подавляющего большинства
континентальных во-
доемов,
как естественных, так и искусственных.
В последнем слу-
чае
развитие экосистем обычно идет по
схеме вторичной сукцес-
сии,
отличается быстрой сменой последовательных
серий и дости-
жением
кульминации на протяжении одного
вегетационного сезона.
Такая
картина, в частности, наблюдается в
повторно заливаемых
рыбоводных
прудах, в рисовых чеках. В естественных
условиях
вторичная
сукцессия типична для некоторых
заливаемых в поло-
водье
придаточных водоемов рек. Скорость
развития экосистем
при
вторичной сукцессии связана с тем, что
в ложе водоема после
его
высыхания остается много зачатков
растений и животных, а
также
продуктов метаболизма существовавшего
здесь биоценоза.
После
нового заполнения ложа в образовавшемся
водоеме очень
быстро
развивается богатое по видовому составу
население, кото-
362V
![]()
рое
при небольшом объеме водной массы
начинает энергично воз-
действовать
на биотоп, преобразуя его в соответствии
с общим
направлением
автотрофной сукцессии.
В
длительно существующих естественных
водоемах автотроф-
ная
сукцессия наиболее выражена в небольших
озерах, в част-
ности
тех, которые в силу климатических,
гидрологических, оро-
графических
и других особенностей благоприятны
для жизни.
Хорошая
прогреваемость воды, поступление с
водосбора большого
количества
биогенов и готового органического
вещества способст-
вуют
нарастанию биомассы и повышению
функциональной роли
биотического
компонента в экосистеме. Озерный сток
происходит
обычно
за счет водной массы эпилимниона,
сравнительно бедной
биогенами,
что способствует их накоплению в
водоеме и ускоряет
сукцессию
экосистемы. В этом же направлении
действуют все
факторы
эвтрофикации озер, связанные с
хозяйственной деятель-
ностью
человека.
Крайне
слабо или вовсе не выражена сукцессия
в реках. Ос-
новная
масса продуктов метаболизма речных
биоценозов уносится
течением
и не изменяет исходных биотопов. Те
небольшие измене-
ния,
которые происходят в экосистеме на
протяжении вегетацион-
ного
периода (например, некоторое заиление
плесов), уничто-
жаются
во время паводка, когда русло промывается,
и река возв-
ращается
в экологически исходное состояние.
Следовательно,
отсутствует
основное условие сукцессии, когда одни
популяции,
модифицируя
среду, создают условия для вхождения
в экосистему
новых
видов. На примере реки видно, как внешние
по отношению
к
экосистеме воздействия препятствуют
ее развитию.
Для
Мирового океана из-за сильной
перемешиваемости воды
сукцессии
не характерны, хотя часто имеют место
процессы, внеш-
не
напоминающие их. Так, при заилении
жестких грунтов их
биоценоз
из кустистых и приподнимающихся форм
заменяется
сообществом
роющих видов. Но в данном случае заиление
— ре-
зультат
не жизнедеятельности биоценоза, а
изменений условий
аккумуляции
осадков, т. е. не является экосистемным
процессом
(Беклемишев,
1973). В ином аспекте, по-видимому, следует
рас-
сматривать
изменение биоценозов по мере удаления
от места
поднятия
глубинных вод, богатых биогенами.
Высокая концентра-
ция
биогенов, обусловливающая в этих местах
формирование
обильного
фитопланктона, служащего пищей различным
его потре-
бителям,
не является результатом жизнедеятельности
биоценоза.
Однако
с удалением от места подъема глубинных
вод количество
биогенов
вследствие потребления водорослями
постепенно сни-
жается.
Соответственно наблюдается направленная
трансформа-
ция
биоценозов, обусловленная изменением
биотопа под влиянием
биотического
компонента.
Гетеротрофная
сукцессия. В некоторых случаях биоценозы
мо-
гут
известное время существовать за счет
утилизации готового
органического
вещества, образованного другими
гидробиоценозами
или
попадающего в водоемы в результате их
загрязнения различ-
363
ными
стоками. Когда, например, новые
водохранилища затопляют
плодородные
земли, богатые растительностью, то
первые 2—3 года
количество
гетеротрофов в водоеме оказывается
непропорциональ-
но
высоким. На грунте в огромных количествах
развиваются де-
тритоядные
личинки насекомых, в частности хирономид.
За счет
массового
размножения бактерий, перерабатывающих
готовое ор-
ганическое
вещество, в водной толще водохранилищ
появляется
богатая
фауна, представленная простейшими,
коловратками и
рачками.
Значительную роль в их питании играет
органическое
вещество,
образуемое продуцентами, однако
потребление пищи
заметно
превосходит ее новообразование. Через
3—4 года био-
масса
бентоса начинает заметно сокращаться,
и постепенно расход
энергии
приходит в соответствие с ее поступлением.
Потребление
кислорода
биоценозами снижается, отношение
рассеиваемой энер-
гии
к аккумулируемой фотосинтетиками
постепенно уменьшается,
но
остается заметно больше единицы. Это
объясняется значитель-
ным
поступлением в водохранилища аллохтонного
органического
вещества,
за счет которого в той или иной мере
существуют многие
гидробионты.
Особенно
резко гетеротрофная сукцессия
проявляется в сточ-
ных
водах с большим количеством органического
вещества. На-
пример,
канализационные воды, выпускаемые для
очистки в спе-
циальные
водоемы, вначале полностью лишены
автотрофов. Их
население
практически представлено бактериями
и очень немно-
гими
животными с анаэробным дыханием или
потребляющими
кислород
воздуха (например, личинка мухи
Ег1з(аИз).
В дальней-
шем
по мере минерализации органических
веществ и появления в
воде
следов кислорода в водоеме, богатом
пищей, появляются
различные
эвриоксибионтные формы (черви ТиЬЦех
IиЫ(ех,
личин-
ки
комара СЫгопотиз,
инфузории Рагатаесшт
рШппит
и др.),
и
видовой состав биоценоза обогащается.
Число видов пока еще
остается
малым, но каждый из них представлен
большим количе-
ством
особей, поэтому биомасса сообщества
велика. При даль-
нейшей
минерализации органики в биоценозе
появляются первые
автотрофы
(в первую очередь синезеленые ОзсПШопа
и Ркоггт-
ёит),
но их роль пока очень невелика. Процессы
рассеяния энер-
гии
— единственные до появления автотрофов
— начинают сопро-
вождаться
ее аккумуляцией, но потребление
кислорода еще резко
преобладает
над его выделением. Фауна водоема
обогащается за
счет
появления форм, более требовательных
к кислороду. С даль-
нейшим
уменьшением концентрации органических
веществ, мине-
рализуемых
гетеротрофами, условия дыхания
становятся более
благоприятными
и соответственно все большее число
видов входит
в
состав биоценозов. Среди автотрофов
появляются диатомовые,
зеленые,
протококковые и, наконец, высшие
растения. Производ-
ство
кислорода увеличивается, отношение
рассеиваемой энергии и
аккумулируемой
снижается. Биомасса биоценоза уменьшается
за
счет
снижения численности массовых форм.
Информационная
емкость
системы возрастает вследствие увеличения
числа видов,
364
сглаживания
их количественной представленности
и усложнения
пищевой
структуры биоценоза.
Процессы,
более или менее сходные с описанными,
наблюдаются
в
водоемах, в частности в реках, в местах
их интенсивного за-
грязнения
с высоким содержанием органических
веществ. В этом
случае
отдельные стадии гетеротрофной
сукцессии оказываются
локализованными
пространственно, существуя параллельно
и поз-
воляя
сразу наблюдать всю последовательность
сериальных из-
менений
сообщества. У места спуска в водоем
больших количеств
органических
веществ население воды представлено
практически
гетеротрофными
бактериями. С продвижением вниз по
течению
реки
соответственно степени минерализации
сбрасываемых орга-
нических
веществ и улучшению условий дыхания
население водое-
ма
становится разнообразнее. Все в большем
количестве разви-
ваются
автотрофы, их роль в биоценозе повышается
и в случае
полной
очистки сточных вод эвтрофное состояние
системы сме-
няется
олиготрофным.
Флуктуация
и трансформация экосистем. Флуктуации
экосистем
в
наибольшей степени связаны с чередованием
сезонов года, в
меньшей
— с максимумами солнечной активности
и некоторыми
другими
циклическими явлениями. Характер
последовательных из-
менений
экосистем на протяжении года позволил
В. Г. Богорову
(1939)
сформулировать представление о
биологических сезонах в
Мировом
океане. По аналогии с временами года
В. Г. Богоров
выделяет
биологические весну, лето, осень и
зиму.
Биологическая
весна характеризуется обилием
фитопланктона,
но
количество зоопланктона еще не достигает
максимума. Отно-
шение
биомассы фитопланктона к биомассе
зоопланктона — так
называемый
сезонный
показатель
— выражается в это время очень
высокой
величиной, лежащей в интервале от 10 до
100. В зооплан-
ктоне
в большом количестве находятся яйца
и личинки бентосных
животных.
Среди водорослей большинство составляют
виды,
нуждающиеся
в сравнительно высоких концентрациях
питательных
солей.
Гидрологически биологическая весна
характеризуется уве-
личением
продолжительности и интенсивности
солнечного освеще-
ния,
началом прогревания воды, часто
наблюдающимся опресне-
нием
ее поверхностного слоя, максимумом
биогенов, а в полярных
морях
— и началом освобождения от сплошных
льдов.
Биологическое
лето связано с наступлением
максимального
освещения,
температура воды повышается, количество
биогенов
снижается.
Оно характеризуется резким уменьшением
сезонного
показателя,
который становится равным или меньшим
единицы.
Биомасса
зоопланктона достигает максимума, из
него исчезают
весенние
и появляются летние личинки. Размеры
планктонных
организмов
уменьшаются, среди водорослей руководящая
роль
переходит
к формам, менее требовательным к
количеству пита-
тельных
солей.
Биологическая
осень начинается при максимальных
годовых
температурах,
которые затем понижаются. Заметно
уменьшается
365
длительность
и интенсивность солнечной радиации.
Количество
питательных
солей вначале несколько повышается
за счет разло-
жения
весеннего и летнего планктона, а затем
снова падает. В поляр-
ных
морях осень в биологическом отношении
характеризуется
уменьшением
концентрации фитопланктона, в умеренных
и юж-
ных—
ее вторым максимумом, заметно более
слабым, чем весной.
Сезонный
показатель в полярных морях меньше
единицы или
равен
ей, в умеренных и южных — колеблется
от 1 до 10. В планк-
тоне
появляются наиболее теплолюбивые
формы.
Биологическая
зима наступает при нарастающем
похолодании,
поверхность
полярных морей покрывается льдом,
интенсивность
освещения
наименьшая, количество биогенов
непрерывно нара-
стает
в результате разложения отмирающего
планктона и подъема
богатых
питательными солями глубинных вод к
поверхности.
В
биологическом отношении зима
характеризуется минимумом
планктона,
сезонный показатель меньше единицы.
Различные виды
водорослей
часто находятся в стадии покоя или
зимующих кле-
ток,
в зоопланктоне главное значение имеют
взрослые животные
или
их зимующие стадии, новых генераций
не образуется.
В
водоемах разных широт одни и те же
биологические сезоны
имеют
неодинаковую продолжительность и
приходятся на разные
месяцы.
Вегетационный период, соответствующий
биологическим
весне,
лету и осени, с 12 месяцев в экваториальных
морях сокра-
щается
до одного-двух месяцев в полярных,
биологической зимы
нет
в экваториальных водах, а в полярных
она достигает 10—-11
месяцев.
С продвижением к полюсам срок наступления
биологиче-
ской
весны запаздывает: в тропических морях
она начинается в
зимние
месяцы, в полярных — в августе.
Продолжительность био-
логического
лета в тропических водах — семь месяцев,
в поляр-
ных—
от одного до двух месяцев. С продвижением
на север и
юг
биологическая осень наступает все
раньше, ее продолжитель-
ность
сокращается и в высоких широтах
становится мимолетной.
Для
сезонных изменений населения пелагиали
характерны не
только
определенные сдвиги в видовом составе
и количественном
соотношении
разных форм. Одновременно четко
прослеживаются
изменения
в возрастной и размерной структуре
популяций, в тро-
фической
структуре биоценозов. Например, в
тропической Атлан-
тике
средняя масса копепод весной меньше,
чем осенью, что
связано
с годовым циклом их развития и повышением
численно-
сти
молодых стадий весной. Максимум развития
продуцентов,
здесь
наблюдается в феврале — апреле,
консументов первого по-
рядка
— в апреле — августе, консументов
второго порядка — в ав-
густе—
декабре. Такой же сдвиг во времени
роста популяций
последовательных
трофических уровней представляет
собой адап-
тацию
потребителей к утилизации своей
кормовой базы.
Весьма
значительные изменения водных экосистем
наблюдаются
в
результате антропогенных воздействий
на водоемы. Зарегулиро-
вание
стока рек ведет к перестройке речных
экосистем в водо-
хранилищные,
значительные отъемы воды на орошение
и другие
366