ПОПУЛЯЦИИ
ГИДРОБИОНТОВ
И
ГИДРОБИОЦЕНОЗЫ
СТРУКТУРА
И ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ
ПОПУЛЯЦИИ
ГИДРОБИОНТОВ
Под
популяциями понимают группу
взаимодействующих друг
с
другом особей одного вида, занимающую
часть ареала этого вида,
обладающую
определенной структурой, способностью
к самовос-
лроизводству
и адаптивной реакцией на изменение
внешней среды.
Это
открытая саморегулирующаяся система,
элементарная форма
существования
вида в тех или иных конкретных условиях
среды. Ес-
ли
условия жизни в пределах- ареала вида
однородны, особи, обита-
ющие
в его отдельных участках, не образуют
популяций, отлича-
ющихся
друг от друга по экологическим и
морфофизиологическим
признакам.
Когда условия жизни распределены
дискретно и в пре-
делах
этих дискретностей относительно
стабильны, вид, как прави-
ло,
распадается на ряд различающихся между
собой популяций.
При
плавной смене биотопических условий
возникает так называ-
емая
клинальная
изменчивость, при которой различия
между сосед-
ними
популяциями малы, но хорошо выражены
между занимающи-
ми
крайние положения.
Виды,
представленные несколькими хорошо
различающимися
между
собой популяциями, называют полиморфными
или полити-
пическими
в отличие от мономорфных,
или монотипических,
не рас-
падающихся
на обособленные группы. Моно- и политипия
— приспо-
собления
вида к менее и более разнобразным
условиям. Чем боль-
ше
выражен биологический полиморфизм,
чем разнообразнее осо-
би,
входящие в популяции, тем легче они
приспосабливаются к цик-
лическим
и внезапным изменениям условий обитания.
По этой при-
чине
виды умеренных широт, живущие в лабильных
абиотических
и
биотических условиях, обладают более
сложной структурой, чем
тропические;
виды, обитающие на широкой акватории,
более поли-
морфны,
чем имеющие узкий ареал.
Биологическое
значение популяций состоит в наиболее
рацио-
нальном
использовании жизненных ресурсов, в
первую очередь
энергетических.
Оптимизация энергетики — основная
тенденция в
становлении
и развитии популяционной структуры.
Среда, как об-
разно
выражается Н. П. Наумов (1977), служит
организмам плац-
дармом
для накопления и передачи взаимной
информации, т. е. их
237Глава 8
биологическим
сигнальным полем,
способствующим групповой кон-
солидации
членов популяции.
В
ряде случаев очень трудно или невозможно
выявить отдельные
популяции
у многих видов планктонных и нейстонных
организмов.
Перенос
течениями на большие расстояния,
особенно в морях и ре-
ках,
исключает существование многих
планктонтов и нейстонтов в
постоянных
географических координатах, ведет к
интенсивному пе-
ремешиванию
особей в пределах ареала вида и
препятствует обра-
зованию
самостоятельных популяций. Сходное
значение имеет пе-
ренос
планктонтов, а также их зачатков
воздушными течениями и
животными
из одного водоема в другой. В связи с
отсутствием в пре-
делах
вида локальных популяций у многих
планктонтов нет и гео-
графической
изменчивости. Иногда среди гидробионтов
встречаются
совокупности
особей одного вида, не воспроизводящие
себя и дли-
тельное
время существующие в данном биотопе
за счет поступле-
ния
новых особей извне. Такие совокупности
представляют собой
псевдопопуляции.
Слагаясь
из организмов, популяции как биологические
системы
характеризуются
новыми качествами, не вытекающими
непосред-
ственно
из свойств отдельных особей. К ним
относятся величина
популяций,
их плотность, дисперсия, половая и
возрастная структу-
ра,
межорганизменные взаимодействия,
рождаемость, рост числен-
ности
и биомассы, смертность и некоторые
другие характеристики,
неприложимые
к отдельным организмам.
Указанные
параметрические характеристики
приспособительно
меняются
(в диапазоне видовых возможностей) в
соответствии
с
периодической или эпизодической сменой
условий существования,
обеспечивая
в конечном счете саморазвитие системы
в направлении
максимизации
ее роли в биосфере.
Из
перечисленных свойств популяций особое
значение для чело-
века
имеет воспроизводство ими своей
биомассы, т. е. образование
органического
вещества в форме тех или иных организмов.
Это
свойство
называется продуктивностью
популяций, а образуемое ими
органическое
вещество — продукцией.
В зависимости от того, в ка-
ких
организмах и формах накапливается
органическое вещество,
продукция
популяций может иметь для человека
совершенно раз-
ное
практическое значение (например,
организмы полезные, беспо-
лезные
или вредные). Темп, эффективность и
энергетические осо-
бенности
продуцирования зависят от специфики
популяций и усло-
вий
их существования.
Хотя
каждое из свойств популяции оценивается
средними стати-
стическими
величинами со значительным рассеянием
значений от-
дельных
параметров, каждая популяция уникальна
по своему эко-
логическому
облику. Как все свойства организмов
выработались в
процессе
эволюции и имеют адаптивное значение,
так и все пара-
метры
популяции отражают ее приспособленность
к существованию
в
тех или иных конкретных условиях. В
этом аспекте и следует оце-
нивать
все свойства популяций и их реакции
на те или иные внеш-
ние
воздействия.
238
Приспособительные
возможности популяции неизмеримо
выше,
чем
у образующих ее отдельных особей.
Популяция определяет свою
судьбу,
«дирижируя», по выражению акад. С. С.
Шварца, физиоло-
гическим
состоянием особей и даже протекающими
в них цитогене-
тическими
процессами. В условиях повышенной
смертности в попу-
ляции
возникает сигнал «нас становится
слишком мало», который
приводит
к кардинальным изменениям физиологии
всех членов по-
пуляции
и ее структуры, направленным к единой
цели: мобилизо-
вать
все экологические резервы в интересах
самосохранения. В об-
ратном
случае в популяции включаются адаптации,
предупрежда-
ющие
развитие опасного для нее процесса
перенаселения.
Структура
популяций
Популяция
не гомогенна по качеству составляющих
ее элементов
(организмов)
и их распределению в пространстве.
Образующие ее
особи
различаются по возрасту, полу,
половозрелости и другим био-
логическим
показателям, причем соотношение групп
разного состоя-
ния
активно поддерживается и регулируется
соответствующими го-
меостатическими
механизмами. Поэтому можно говорить
о возраст-
ной,
половой, генеративной или иной структуре
популяций
применительно
к их качественному составу. Помимо
этого, структу-
ра
популяций характеризуется их величиной,
плотностью (концен-
трацией)
и особенностями размещения в пространстве
(хорологи-
ческая
структура).
Необычайно
характерна и биологически важна
морфофизиоло-
гическая
разнокачественность особей одного
возраста и пола в
каждый
из моментов существования популяций.
Все параметры
структуры
динамичны, сопряжены между собой и
обеспечивают
каждым
конкретным сочетанием оптимальный
вариант состояния
популяций
применительно к меняющимся условиям
среды.
Величина
и плотность. Величина популяций
определяется чис-
лом
и суммарной массой (биомассой) образующих
ее особей без-
•относительно
к величине занимаемого ими пространства.
В одних
•случаях
популяции имеют огромную численность
и сравнительно
малую
биомассу (бактерии, водоросли,
простейшие), в других на-
блюдается
иная картина (крупные моллюски, рыбы,
млекопита-
ющие)
. Соотношение между численностью и
биомассой популяции —
один
из ее характерных признаков. Даже
популяции одного вида,
обитающие
в разных водоемах или их отдельных
участках, иногда
значительно
разнятся между собой по соотношению
численности и
биомассы
особей, что опять-таки характеризует
состояние популя-
ции.
Крайне
различна величина популяций данного
вида в отдельных
водоемах
или их участках. Как правило, популяции
гидробионтов,
обитающие
в водоемах или их участках с большой
акваторией,
многочисленнее,
так как имеют больше жизненных ресурсов.
С уве-
личением
популяций меняются их многие качества,
в частности ус-
тойчивость
существования и способность расширять
свой ареал.
239
Чем
больше особей в популяции, тем богаче
ее генофонд и шире
диапазон
каждого фактора, в пределах которого
могут существовать
те
или иные особи. Поэтому резкие изменения
среды, не всегда пе-
реносимые
малой популяцией, легче выдерживаются
большой.
Часть
ее особей, оказавшихся за пределами
своей экологической
валентности,
погибает, но другие, с иными параметрами
толерант-
ности,
выживают, и популяция сохраняется.
Характерно, что в низ-
ких
широтах, где условия существования
стабильнее, популяции
малочисленнее,
чем в умеренных и полярных зонах.
Возможно, что
именно
благодаря своей малочисленности
популяции гидробионтов,
обитающие
в тропических морях, легче проникают
в континенталь-
ные
водоемы, чем высокоширотные формы.
Последние адаптирова-
ны
к существованию в больших популяциях
и потому не могут про-
никать
в водоемы, недостаточные для поддержания
количества осо-
бей
на требуемом высоком уровне из-за
ограниченности их ресурс-
ной
базы (Численно, 1964).
Весьма
характерный параметр состояния
популяции — ее плот-
ность,
под которой понимают отношение
количества организмов к
единице
занимаемого ими двух- или трехмерного
пространства. Ко-
личество
организмов может быть выражено числом
особей, их био-
массой
или в энергетических единицах (например,
в джоулях).
Наиболее
полное представление о плотности
популяции дает харак-
теристика
ее всеми перечисленными показателями.
Численность
особей
характеризует их среднюю удаленность
друг от друга, био-
масса—
концентрацию живого вещества,
энергоемкость — термо-
динамические
особенности.
Плотность
популяции определяется равновесием
между потен-
циалом
размножения и ресурсными возможностями
среды — ем-
костью
среды.
Последняя в свою очередь обусловливается
скоростью
возобновления
требуемых ресурсов, величиной их
продукции в еди-
ницу
времени. Этот параметр — не константа,
так как, в частности,
в
сильной мере зависит от характера
эксплуатации возобновля-
ющихся
ресурсов. Например, продукция дафний
как возобновимого
кормового
ресурса рыб может оказаться минимальной
из-за чрез-
мерного
выедания особей. Понятно, что на скорость
возобновления
ресурсов
помимо формы эксплуатации влияет ряд
других биотиче-
ских
и абиотических факторов.
Плотность
популяции определяет величину ее
суммарного обме-
на,
отнесенного к единице занимаемого
пространства. С уменьше-
нием
размера организмов интенсивность их
обмена возрастает. По-
этому
чем они мельче, тем при меньшей их общей
биомассе дости-
гается
определенная величина суммарного
метаболизма, играющая
ту
или иную роль в ограничении плотности
популяций. Это положе-
ние
подтверждается, в частности, при
выращивании различных
гидробионтов.
Например, в оптимальных условиях число
особей
крупных
ОарНта
тадпа,
развивающихся и живущих одновременно
в
1 м3
«оды, в несколько раз ниже, чем мелких
Мота
гесИго$1пз,
а
биомасса выше (Шпет, 1962). Такое соотношение
прослежено и
при
выращивании разноразмерных личинок
хирономид. На рис. 41
240
к1000
200
ъ
иж'1
I
[100
^
ь
I
[50
^ •
О
Личинки.
Мальки Сеголетки Двухлетки
масса25мг
тссагг тссагог жса280г
350
300
250
§
200
150
50
Сз
Сй
Число
■
Масса
Рис.
41. Нормы посадки личинок, мальков,
сеголеток и
двухлеток
карпа на 1 м3
воды пруда
показано,
как снижается предельно допустимая
биомасса карпов в
случае
зарыбления прудов молодью убывающего
размера.
Плотность
популяции представляет собой один из
важнейших
параметров
ее существования. От плотности зависят
дыхание, пита-
ние,
размножение и многие другие функции
отдельных особей по-
пуляции.
Например, скорость фильтрации моллюсков
5ркаепит
сотеит
с увеличением их числа в подопытных
сосудах сначала воз-
растает,
потом снижается (Алимов, 1967):
Число
моллюсков в сосуде 1 5 10 15 20
Скорость
фильтрации, мл/ч ....
3,4 6,9 7,5 5,2 3,8
Рачки
РагсеИо
всаЪег
за 30 дней выращивания в группах по
экз.
давали индивидуальный прирост 7,3 г, а
в группах по
10
экз.— 8,1 г. Молодь мурманской сельди
совсем не питается, ког-
да
рыбки содержатся поодиночке, и через
некоторое время погиба-
ет
от истощения. В группах из 5 особей
молодь начинала потреблять
корм
через 3—4 дня после посадки в аквариум,
а когда сажалась
по
20 экз.— на следующий день. Молодь
азовского леща в стае
питается
интенсивнее, чем в одиночку, а взрослые
рыбы, наобо-
рот,
потребляют корма больше, когда находятся
вне стаи. У ряда
рыб
и беспозвоночных отмечена значительная
зависимость интен-
сивности
дыхания от чисел особей, помещаемых
в одинаковые по
размерам
сосуды. Интересно, что групповой эффект
иногда может
возникать
только на основе зрительного восприятия
особей. Так,
золотые
рыбки снижали потребление кислорода,
когда содержались
поодиночке,
но видели свое отражение в зеркале.
241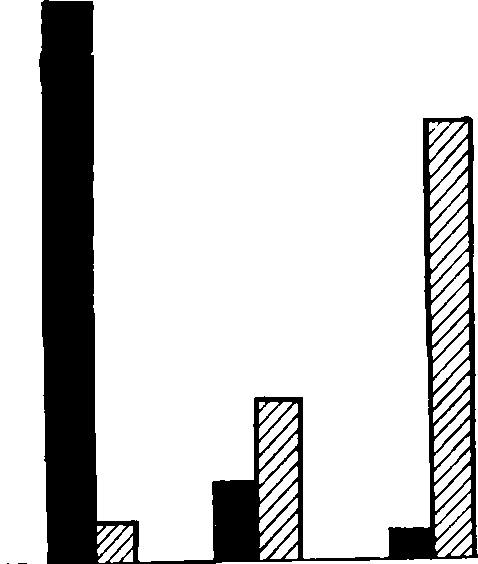
.
Чрезмерная плотность популяции ухудшает
условия ее существо-
вания,
снижая обеспеченность особей пищей и
другими средствами
к
жизни. Характерно, что у морских ежей
ЕсМпотеЬга
таНгаеь
с
повышением плотности популяции
увеличиваются относительные
размеры
аристотелева фонаря — органа добывания
пищи (В1аск е!
.а11.,
1982).
Отрицательно
влияет на популяцию и недостаточная
плотность
ее,
затрудняющая отыскание особей
противоположного пола, созда-
ние
нужной концентрации половых продуктов
в воде во время раз-
множения,
выработку защитных свойств популяции
и др. Для каж-
дого
вида существует оптимальная плотность
популяций, которая
приспособительно
меняется в зависимости от конкретных
состояний
среды,
т. е. является адаптивным признаком.
Имеется
ряд механизмов поддержания плотности
популяций на
нужном
уровне. Главный из них — саморегуляция
численности по
принципу
обратной связи с количеством жизненных
ресурсов, в ча-
стности
пищи. Когда ее становится меньше, темп
роста особей сни-
жается,
смертность возрастает, половая зрелость
наступает позже,
пополнение
популяции снижается, плотность падает.
Улучшение
условий
существования сопровождается изменениями
противопо-
ложного
характера, и плотность популяций
возрастает до определен-
ного
предела, превышение которого снова
ведет к уменьшению кон-
центрации
особей.
Наиболее
часто плотность регулируется изменением
темпа раз-
множения.
Например, в переуплотненных популяциях
дафнии обра-
зуют
покоящиеся яйца, и плотность популяции
животных снижает-
ся.
С повышением плотности популяции
обычно снижается плодо-
витость
животных. Так, с увеличением концентрации
Оаркта
ри1ех-
с
1 до 4, 8, 16 и 32 экз./мл среднее число
потомков на одну самку
за
один день снижается с 4 до 2,8; 1,8; 0,4 и
0,1. Имеются данные о
том,
что плотность популяций водорослей
может регулироваться час-
тотой
столкновения клеток. Так, культура
Те1гакутепа
руп}огт18
перестает
расти, достигая плотности 0,5 млн. кл./мл.
Если в нее
вносились
дутые стеклянные шарики, по размеру
равные клеткам,
число
последних сокращалось так, что
концентрация клеток и ша-
риков
не превышала 0,5 млн. кл./мл; после удаления
шариков куль-
тура
снова достигала начальной плотности
(5аИоЬ, Аза!, 1982).
С
увеличением плотности популяций все
более частой и массо-
вой
становится эмиграция особей в результате
активных и пассив-
ных
перемещений. У малоподвижных бентосных
форм наблюдается
подъем
особей в толщу воды и перенос их
течениями иногда на
очень
большие расстояния. Периодически
поднимаются в толщу
боды
и
сносятся вниз по течению многие
представители речного бен-
тоса.
Излишнее
увеличение плотности в популяциях
многих данных
животных,
особенно морских, предупреждается
образованием пе-
лагических
личинок, долго находящихся в толще
воды и переноси-
мых
течениями иногда на сотни и тысячи
километров. Это особен-
но
характерно для малоподвижных моллюсков,
иглокожих и при-
242
крепленных
форм (усоногих ра- §
ков,
кораллов, морских лилий
и
др.)-
Один
из важных механиз- ^
мов
регуляции плотности попу-
ляций
— выделение метаболи-
тов,
воздействующих на рост и
развитие
особей. На рыбах по-
казано,
что в присутствии соб-
ственных
видоспецифических
экзометаболитов,
когда их кон-
центрация
достигает некоторой
критической,
большая часть
особей
замедляет рост, мень-
шая—
сохраняет или даже ус-
коряет
его. В результате сум-
марный
прирост особей замед-
ляется
и исходная группа од-
норазмерных
рыб резко диффе-
ренцируется
по размерному со-
ставу
(рис. 42).
В
сильнейшей мере плотность популяции
гидробионтов, особен-
но
бактерий, водорослей и беспозвоночных,
зависит от выедания их
другими
организмами. С повышением плотности
популяций пере-
численных
групп гидробионтов возрастает кормовая
ценность обра-
зуемых
ими пастбищ и соответственно увеличивается
приток по-
требителей,
снижающих концентрацию поедаемых
организмов. Уход
потребителей
с оскудевших пастбищ создает благоприятные
усло-
вия
для повышения плотности популяций
кормовых организмов.
Сходное
регулирующее воздействие оказывает
паразитарный фак-
тор.
Чем выше численность особей и больше
контакт их друг с дру-
гом,
тем эффективнее циркуляция паразитов
и тем сильнее они воз-
действуют
на численность хозяев. Плотность
популяций промысло-
вых
гидробионтов во многих случаях
определяется интенсивностью
их
вылова.
Хорологическая
структура. В пространстве, занимаемом
попу-
ляцией,
распределение особей может быть
беспорядочным,
однооб-
разным
и пятнистым.
При беспорядочном, или случайном,
распре-
делении
вероятность встретить особь в каждой
из точек одинакова.
Такой
тип распределения встречается
сравнительно редко, в част-
ности,
когда среда однообразна и у организмов
нет тенденции к
агрегации
или дезагрегации. Если среда однообразна
и особи стре-
мятся
селиться на каком-то удалении друг от
друга, наблюдается
равномерное
распределение.
Для
гидробионтов наиболее характерно
пятнистое распределе-
ние,
которое в свою очередь может быть
беспорядочно-кучным, од-
нообразно-кучным
и агрегатно-кучным. В первом случае
особи в
пятне
располагаются случайно, во втором —
на сходных расстояни-
ях
друг от друга, в третьем — группами.
Пятнистое распределение
сз
*
I
’
§
\
!<
§
-/
§
г
_1
50
ПО
■* КО
_
Концентрация
видоспецифических
экзометаболитов,
уел. ед.
Рис.
42. Удельная скорость роста моло-
ди
карпа и белого амура при разной
кон-
центрации
собственных видоспецифиче-
ских
экзометаболитов:
/
— карп, 2
—
белый амур
243I..
— 1
имеет
место, если условия среды, наиболее
благоприятные для су-
ществования
особей, распределены мозаично, что
чаще всего и на-
блюдается
в водоемах, особенно в бентали. Пятнистый
характер
распределения
организмов в основном определяется
взаимоотноше-
ниями
между особями.
Одним
из показателей особенностей распределения
служит от-
ношение
дисперсии к среднему значению,
вычисленному по количе-
ству
оробей на ряде пробных площадок. Если
через Х0
обозначить
среднюю
численность особей на единицу площади
или объема ис-
следуемого
пространства (на основе учета количества
организмов
в
п
пробах), то дисперсию (52)
можно найти по формуле
П
82='^(хд—х1)2-(п—
1), где х
— численность особей в 1-той
про-
/-1
«бе.
Если отношение 52/лг0<1,
распределение равномерное, >1
—
пятнистое,
равно 1 — случайное.
Рассредоточиваясь,
особи проявляют свою активность на
неко-
торой
площади. Если эту площадь организм
защищает от вторже-
ния
других особей, ее называют «территорией»,
а само владение —
территориальностью.
Территориальность, представляющая
собой
упорядочение
использования мест обитания, наиболее
часто про-
слеживается
у бентосных форм, особенно у высших
раков, мол-
люсков,
иглокожих. Например, крупный литоральный
моллюск
ЬоШа
%щап1еа,
питающийся водорослевой пленкой,
охраняет аква-
торию
в 300—900 см2,
являющуюся для него пастбищем. С
увели-
чением
размеров моллюсков и снижением толщины
пленки охраня-
емая
территория возрастает. Если удалить
крупных моллюсков, во-
доросли
быстро уничтожаются другими животными.
С кормовой ба-
зой
связана и территориальность у
широкопалого рака.
Обитающие
в трубках амфиподы ЕпсМНотиз
ЪгавШепвьз
избе-
гают
селиться- вблизи друг друга и защищают
свою кормовую тер-
риторию
от новых вселенцев; подобная картина
описана для мол-
люсков
ТеШпа
1епш8,
крабов Пса
ридьШог
и V.
рицпас1а,
для лан-
густа
Iавиз
Шапйеи
Офиура ОрЫосота
8Со1орепйппа
прогоняет со
своего
кормового участка других офиур. Морские
блюдечки Ра1е11а
еотрге88а
живут только на крупных водорослях по
одному на тал-
ломе,
не допуская на территорию других особей
своего вида. Дру-
гой
представитель морских блюдечек ЬоШа
ц1цап1еа,
прогоняя при-
шельцев,
пытается оторвать их от субстрата или
толчками вынуж-
дает
к удалению, не преследуя отступающих
(АУп^Ы, 1982). Самцы
гавайского
краба не допускают постройки нор других
особей в ра-
диусе
70 см от своей норы, отталкивая прищельцев
клешнями.
Нередко
в пределах одной популяции возникают
различные
скопления
и группировки особей. Они образуются
в соответствии
с
принципом «биологики», под которой в
бионике понимают опреде-
ленную
организацию биологических элементов,
позволяющую их
совокупности
совершать то, чего каждый из них не мог
бы сделать
в
отдельности. Нахождение в группах в
какой-то степени невыгодно
особям
(усиление конкуренции и др.), но
преимущества доминиру-
244
ют
над негативными моментами, и потому
скопления повышают вы-
живаемость
популяции в целом. Нередко в группах
наблюдается
•снижение
смертности при нападении других
организмов или ухуд-
шении
абиотических условий (кондиционирование
среды). Напри-
мер,
при совместном содержании рыбы
выдерживают большие дозы
токсикантов,
вредное действие которых нейтрализуется
выделени-
ем
значительных количеств слизи и других
экскретов. Находясь в
стае,
гидробионты успешнее обороняются от
хищников и эффектив-
нее
нападают на жертву. Часто объединение
в группы бывает вре-
менным,
и степень агрегации особей маневренно
меняется в соот-
ветствии
с тактикой существования популяций,
настраивающихся
на
среду.
Возрастная
структура. Соотношение в популяции
особей раз-
ного
возраста, будучи приспособительным
свойством видов, меня-
ется
в зависимости от состояния среды. В
благоприятных условиях,
жогда
наблюдается прирост популяций, в них
относительно высока
численность
молодых особей. В популяциях, величина
которых не
меняется,
относительное количество молоди ниже
и еще меньше оно
в
популяциях, сокращающихся вследствие
падения рождаемости.
Для
одних видов характерна многовозрастная
структура, другие
представлены
организмами с короткими жизненными
циклами. Эти
различия
не случайны. Как показал Г. В. Никольский,
для рыб
(и
это справедливо в отношении многих
других водных животных)
многовозрастная
структура отражает приспособленность
популяций
к
устойчивой кормовой базе, слабому
уничтожению взрослых осо-
бей
и лабильным условиям воспроизводства;
животные с корот-
ким
жизненным циклом адаптированы к
противоположным услови-
ям.
Возрастная структура популяции
приспособительно перестраи-
вается
как самонастраивающаяся система через
изменение темпа
роста,
созревания и длительности жизни.
Возрастная
структура популяции циклически
меняется, если об-
разование
новых генераций
(поколений) носит периодический
ха-
рактер,
как это наблюдается в подавляющем
большинстве случаев.
У
многих водорослей новые клетки
образуются преимущественно
днем,
поэтому их популяции в это время суток
содержат больше
молодых
особей, чем ночью. В популяциях,
представленных орга-
низмами
с более длительным жизненным циклом,
периодичность
пополнения
может определяться фазами луны, и тогда
возрастная
структура
перестраивается каждые 14 или 28 дней.
Так меняется она
у
червя ЗрггогЫз
ЬогеаЫз,
моллюсков Масота,
ЬШоппа, МуШиз,
у
креветок АпсЫзИоШез
и ряда других форм, хотя некоторые
внеш-
ние
условия, например штормы, могут нарушать
подобную ритмику.
Крайне
характерны для популяций гидробионтов
сезонные измене-
ния
возрастной структуры. Например, весной
в водоемах появляется
огромное
количество молоди животных, а осенью
и зимой в них
больше
старших возрастных групп. У форм,
размножающихся раз
в
год или реже, возрастная структура
сильно зависит от урожай-
ности
отдельных поколений. На улучшение
условий существования
популяции,
как правило, реагируют изменением
возрастной струк-
245
туры
в сторону повышения роли молодых
особей, на ухудшение —
противоположным
образом. Такое изменение возрастной
структуры
обусловливается
не только возрастанием темпа
новообразования
молоди,
но и повышением их выживаемости в
благоприятных усло-
виях.
Характерные
изменения возрастной структуры
популяций на-
блюдаются
с продвижением к экватору. В низких
широтах преоб-
ладают
популяции с коротким жизненным циклом
особей и неболь-
шим
числом возрастных групп, что позволяет
им выдерживать зна-
чительную
смертность в условиях интенсивного
выедания. В аркти-
ческой
и бореальной зонах популяции отличаются
более разновоз-
растной
структурой, обеспечивающей устойчивость
воспроизвод-
ства
в резко меняющихся условиях внешней
среды. Последнее об-
стоятельство
не имеет существенного значения для
тропических по-
пуляций,
особи которых могут размножаться
круглый год.
Возрастная
структура популяций в сильной мере
определяется
степенью
выедания особей разных возрастов. Как
правило, в боль-
шей
мере уничтожаются мелкие, менее
защищенные особи, в част-
ности
молодь червей, моллюсков, ракообразных,
рыб. Соответствен-
но
этому у перечисленных групп особенно
велика разница в коли-
честве
младших и старших возрастов. У живородящих
червей, мол-
люсков,
ряда рыб и у многих водных млекопитающих
молодые ста-
дии
не так сильно уничтожаются, и возрастная
структура этих
животных
меньше различается по численности
младших и старших
групп.
У промысловых гидробионтов структура
часто определяется
характером
вылова и обычно отличается низкой
относительной чис-
ленностью
крупномерных особей, особенно в условиях
перелова.
Половая
и генеративная структура. Половая
структура популя-
ций
характеризуется соотношением в них
особей разного пола.
С
увеличением количества самок возрастает
темп воспроизводства
популяции,
но снижается жизнестойкость пополнения.
Поэтому в
благоприятных
условиях у некоторых животных резко
преобладают
самки
вплоть до полного исчезновения самцов.
Так, полное отсутст-
вие
их наблюдается летом у многих низших
ракообразных и коло-
враток.
Когда условия ухудшаются, относительная
численность
самцов
в популяциях повышается, и это ведет
к увеличению жиз-
нестойкости
потомства. Равная представленность
самцов и самок,
обеспечивая
удовлетворительный темп пополнения
популяции и
достаточный
уровень ее жизнестойкости, в наилучшей
степени от-
вечает
требованиям перекомбинирования
наследственного матери-
ала
и обогащения генофонда, что резко
повышает выживаемость
популяций
в меняющихся условиях среды.
Соотношение
полов в первую очередь зависит от
наследствен-
ных
свойств вида, но в значительной мере
контролируется внешни-
ми
условиями. Когда, например, молодь
рачков Саттагиз
йиеЪет
выращивали
в условиях длительного светового дня
(16 ч освеще-
ния),
самцов появлялось в 3—12 раз больше,
чем самок, но послед-
ние
резко преобладали (4:1) при коротком
световом дне (8 ч осве-
щения).
В аналогичных опытах с мокрицей
АгтайШШит
ьи1цаге
246
смещение
соотношения полов носило противоположный
характер.
Световой
режим действует только на критическую
фазу, когда пол
еще
не определился.
Существенно
влияет на половую структуру популяций
темпера-
тура.
Например, с ее повышением от 10—12 до
25—28° С количест-
во
самцов в потомстве рачков Масгосус1орз
а1ЫсИиз
возрастает с
до
64,7% (Монаков, 1965). Сходная картина
прослежена у
Асап1Носус1ор8
ьЬгШз
и ряда других веслоногих. У Саттагиз
заИ-
пиз
при температуре 5° С самцов в 5 раз
больше, чем самок, а при
23°
С — в 13 раз меньше; содержание самок
С.
йиеЪет
в период
окончания
созревания ооцитов при температуре
ниже 5° С вызыва-
ет
появление в потомстве только самцов,
а при 7—8° С — только
самок.
Соотношение самцов и самок в культуре
Юаркта
тадпа
сдвигалось
в пользу первых, когда температура
достигала 27° С, и
в
пользу вторых, когда она понижалась
до 11° С, при 8° С имелись
только
самки, при 30° С — только самцы. В
популяции байкальских
рачков
Сус1орз
ко1епз1з
зимой резко преобладают самцы (70—
#0%),
летом и осенью их становится значительно
меньше
(30-60%).
Общеизвестна
смена в неблагоприятных условиях
однополых
популяций
двуполыми у многих низших ракообразных
и коловра-
ток.
У гермафродитных организмов с регулярным
чередованием
мужской
и женской фаз половая структура зависит
от возрастной.
Например,
промысловая креветка РапйаШз
ЬогеаИз
обычно первый
раз
принимает участие в размножении в
качестве самца в возрасте
2,5
года, затем меняет пол и в следующий
сезон размножения от-
кладывает
икру. Креветка 8о1епосе>га
тетЬгапае1а
живет 3 года,
причем
два из них (последние) функционирует
как самка; по этой
причине
самцов в популяции много меньше и они
мельче. Устрицам
Оз1геа
1и1аг1з
до 2-го или 3-го сезона размножения
свойственно
регулярное
чередование мужской и женской фаз,
после чего они
становятся
гермафродитными. Нередко меняют свой
пол Нуйга
аИепиа{а.
У морской звезды Аз1ег1па
дгЬЬоза
в первые 2 года после
наступления
половой зрелости наблюдается
протерандрический
гермафродитизм,
затем происходит оогенез.
Половая
структура может зависеть от избирательного
уничто-
жения
самцов или самок, а у промысловых
животных — от селектив-
ности
лова.
Генеративная
структура популяций определяется
соотношением
особей,
находящихся в префертильной,
фертильной
и постфертиль-
ной
стадиях, т. е. числом особей, еще не
способных размножаться,
размножающихся
и утративших способность к размножению.
Дли-
тельность
каждой стадии сильно варьирует у
представителей раз-
ных
видов. Например, префертильная стадия
у многих поденок,
двукрылых,
у дальневосточных лососей и ряда других
животных
длится
несколько лет, фертильная — несколько
дней или недель,
а
постфертильная отсутствует, поскольку
организмы названных
групп
после размножения погибают. У большинства
моллюсков,
червей,
рыб и водных млекопитающих длительность
фертильной
247
стадии
выше, чем префертильной. Генеративная
и половая структу-
ра
популяций имеют приспособительное
значение, обеспечивая не-
обходимую
эффективность размножения в меняющихся
условиях
среды.
В соответствии с изменениями условий
жизни генеративная
и
половая структура адаптивно
перестраивается, приобретая пара-
метры,
оптимальные для воспроизводства
популяции применитель-
но
к реально складывающейся ситуации.
Разнокачественность
особей. Морфологическая и физиологиче-
ская
разнокачественность особей в популяциях
— одна из важней-
ших
адаптаций к наиболее эффективному
использованию потенци-
альных
жизненных ресурсов. Формула Ч. Дарвина
«наибольшая
сумма
жизни обеспечивается при максимуме
ее разнообразия» рас-
пространяется
не только на видовое богатство, но и
внутривидовое.
Существование
территориальных, морфологических,
физиологиче-
ских,
экологических и других группировок
вида обогащает его свя-
зи
со средой, повышает возможности более
широкого использования
ее
жизненных ресурсов. То же относится к
отдельным популяциям
вида.
Их разнокачественность обусловлена
структурой — возраст-
ной,
половой, генеративной, сезонной и
некоторыми другими состоя-
ниями.
Но даже в пределах каждого состояния
(возраста, пола
и
т. д.) особи в каждый момент отличаются
друг от друга, и сте-
пень
их варьирования по отдельным признакам
носит приспособи-
тельный
характер, являясь особой формой
«настройки» популяции
на
среду в интересах ее максимального
освоения. Например, у осет-
ровых
рыб, нерестящихся на гальке, икринки,
даже выметываемые
одной
самкой, обладают разной клейкостью,
что обеспечивает их
перенос
течением на разные расстояния и
предупреждает образо-
вание
вредных скоплений. Благодаря варьированию
частоты и дли-
ны
жаберных тычинок у сигов увеличивается
ассортимент кормов,
используемых
популяцией, и ее кормовая база
расширяются. Неод-
нородность
по размеру хирономид, выращиваемых из
одной яйце-
кладки
при большой плотности посадки
(Константинов, 1958), по-
зволяет
группам личинок завершать метаморфоз
последовательно,
что
по мере вылета комаров обеспечивает
успешное развитие осо-
бей,
отставших в росте.
Выработанные
в процессе эволюции и наследственно
закреплен-
ные
нормы реакции, определяющие пластичность
организмов, срав-
нительно
узки, так как значительное расширение
эврибионтности,
полезное
для существования в изменчивых условиях,
сопровожда-
ется
снижением эффективности использования
каждой конкретной
ситуации.
Здесь для отдельных организмов возникает
непреодоли-
мое
противоречие, которое снимается в
популяциях за счет разно-
качественности
особей, помогающей использовать
неустойчивые
жизненные
ресурсы через разнообразие индивидуумов,
различие
их
нормы реакции. Вследствие разнокачественности
повышается
устойчивость
популяции, когда при крайних значениях
какого-либо
фактора
отдельные особи окажутся достаточно
резистентными к
ним
и обеспечат сохранение системы в целом.
Будь популяция
особью,
«тиражированной» в тысячах или миллионах
экземпляров,
248
она
погибла бы при первом значительном
изменении среды, выхо-
дящем
за пределы толерантности организмов
с единым генотипом,
с
единой нормой реакции.
Разнокачественность
особей в популяциях динамична и
подвер-
жена
определенным закономерным изменениям,
как это детально
прослежено
на рыбах. Самая общая закономерность
— преимуще-
ственное
изменение биологических показателей
у наиболее много-
численных
(модальных) групп особей, по степени
выраженности
или
количественному значению признаков
и свойств, связанных с
сильно
изменившимися природными факторами.
Например, рез-
кое
ухудшение условий питания популяций
рыб сопровождается
преимущественным
замедлением скорости роста, снижением
упи-
танности
и плодовитости у особей самых
многочисленных по раз-
меру
или возрасту. Это объясняется тем, что
быстрее всего истреб-
ляются
пищевые организмы, наиболее соответствующие
потребно-
стям
многочисленных по тому или иному
признаку рыб (Поляков,
1975).
«Настройка»
популяции на изменение факторов
сводится к то-
му,
что самые многочисленные группы особей
приобретают такие
признаки,
которые обеспечивают эффективное
использование этих
факторов.
Резкие изменения среды модулируют
имевшуюся струк-
туру
— иным становится распределение особей
по классам различ-
ных
признаков, вершина кривой смещается
(положительная или
отрицательная
асимметрия), иногда кривая становится
многовер-
шинной
или прерывается. Происходящие изменения
усиливают
разнокачественность
популяции, расширяя ее
приспособительные
возможности,
и способствуют быстрейшему восстановлению
соот-
ветствия
между размерами популяции и условиями
жизни. Напри-
мер,
с увеличением дифференциации стада
рыб по размеру расши-
ряется
их кормовая база — ассортимент кормов,
доступных особям
разного
размера. Отсюда понятно, что на ухудшение
кормовых (и
других)
условий популяции гидробионтов
реагируют усилением
разнокачественности.
Например, в уплотненных посадках
личинки
хирономид,
выклюнувшиеся из одной яйцекладки,
отличаются по
темпу
роста, и вылет первых комаров начинается
тогда, когда от-
ставшие
в своем развитии личинки находятся
только на второй
стадии;
в благоприятных условиях особи близки
по размеру, и вы-
лет
комаров становится дружным. Изменчивость
раковины мидий
усиливается
у северных границ ареала вследствие
отклонения ус-
ловий
существования от оптимальных.
У
рыб ухудшение кормовых условий
сопровождается увеличени-
ем
амплитуды изменчивости размеров
одновременно выметываемой
икры
(запаса желтка) и размеров одновозрастных
особей стада;
обратная
картина наблюдается с улучшением
условий питания.
Как
показал Г. В. Никольский (1982) на рыбах,
и это справедливо
в
отношении многих других гидробионтов,
морфологическая, физио-
логическая
и экологическая изменчивость особей
подчинена опре-
деленной
вертикальной и широтной поясности.
Внутривидовая из-
менчивость
у высокоширотных форм выше, причем эта
закономер-
249
ность
резче выражена в пресных водоемах,
поскольку в морях ши-
ротные
контрасты условий жизни не столь
различны.
Внутрипопуляционные
отношения
Характерная
особенность внутрипопуляционных
отношений — их
направленность
на повышение устойчивости системы, на
оптимиза-
цию
ее взаимодействия со средой, на повышение
маневренности и
эффективности
использования жизненных ресурсов.
Формы взаимо-
действия
между особями популяции крайне
многогранны. Это пря-
мая
борьба и взаимопомощь, стимуляция и
ингибирование, конку-
ренция,
иерархия и некоторые другие отношения.
Широко распро-
странено
образование стай и различных скоплений,
обеспечивающих:
популяциям
те или иные биологические преимущества.
Некоторые
формы
внутрипопуляционных отношений (прямая
борьба, конку-
ренция
и др.) внешне напоминают межвидовые.
Однако они имеют
совершенно
другой биологический смысл, поскольку
всегда направ-
лены
на процветание вида, эволюционно
выработались в качестве
адаптаций
к существованию популяций в тех или
иных конкретных
условиях.
Все
формы внутрипопуляционных отношений
базируются на ком-
муникации
особей — химической, вибротактильной,
визуальной и др_
Среди
химических средств коммуникации
огромную роль играют
феромоны
— вещества, имеющие сигнальное значение.
Различают
феромоны
стадные, вызывающие скопление особей
(например, у
планарий,
балянусов), половые (нематоды,
ракообразные, рыбы),
тревоги
(моллюски, иглокожие, рыбы), следовые
(маркированиепу-
ти)
и некоторые другие. Важно иметь в виду,
что все формы связи
эффективно
работают в естественной среде, и ее
изменение создает
опасность
появления «шумов» — искажения
информационных сиг-
налов.
Прямая
борьба и взаимопомощь. Случаи
прямой борьбы наблю-
даются
при разных формах агонистического
поведения
(от лат.
адоша
— борьба, противоборство) и при
каннибализме. Адаптивное
значение
последнего неоднозначно. Например,
щука и окунь, не
способные
потреблять планктон и бентос, могут
быть единственны-
ми
представителями' ихтиофауны в некоторых
озерах, существуя
за
счет поедания части своей молоди,
питающейся мелкими живот-
ными.
В значительной мере поедают собственную
молодь Сус1ор&
ЫсизрШаШз.
В рассматриваемых случаях хищники
через посредст-
во
своей молоди осваивают кормовые
ресурсы, которые не могут ис-
пользовать
непосредственно. Каннибализм наблюдается
у морских
звезд
Аз1епаз
гиЬепз,
бокоплавов Саттагиз
ри1ех,
личинок комара
Ргос1айшз
и ряда других животных, когда популяция
не обеспечена
кормом.
В этих случаях каннибализм помогает
популяции выжить
за
счет
снижения своей численности.
Поедание
одних особей другими может иметь место
и в услови-
ях
обеспеченности пищей, как это, например,
наблюдается в плот-
ных
культурах жгутиковых Регапета
IпскорНогит;
попытки загло-
250
тить
представителей своего вида отмечены
в 68% случаев контактов
животных,
причем, как правило, они заканчивались
поеданием бо-
лее
мелких особей, и плотность культуры
резко снижалась (№ск,
1982).
Чрезвычайно
разнообразны формы агонистического
поведения,
которое,
как правило, представляет собой
столкновение особей за
право
обладания каким-то объектом с минимальной
травматизаци-
ей
партнеров (без преследования или
уничтожения побежденного).
Например,
борьба раков-отшельников за обладание
актинией длит-
ся
до тех пор, пока один из противников
не признает своего пора-
жения,
приняв характерную позу «смирения
побежденного» (ло-
жится
на бок). У крабов Р1еитпи$
зауь
борьба за убежище в коло-
нии
мшанок заканчивается, как только один
из противников на-
чинает
удаляться от нее; чем больше разница
в размерах крабов,
тем
меньше актов демонстрации силы
необходимо для завершения
схватки.
Во время борьбы наблюдается нарастание
«силовых при-
емов».
Например, морские ежи ЕсЫпоте1га
1исип1иг
в борьбе за
нору
сначала толкают друг друга, затем, если
этого недостаточно
для
ухода одного из противников, пускают
в ход зубы аристотелева
фонаря;
при равных размерах победителем обычно
оказывается хо-
зяин
норы (ОгйпЬаиш е1 а11., 1978). Борьба
минимальна, когда
агрессор
много сильнее атакуемого. Мелкие
осьминоги часто без
драки
уступают убежище крупным особям,
молодые речные раки
оставляют
добычу, когда приближаются старые.
У
малоподвижных или неподвижных животных
агонистическое
поведение
нередко выражается в умерщвлении
соседних особей сво-
его
вида. Например, многие актинии убивают
близсидящих особей
нематоцистами,
густо расположенными на особых щупальцах
—
акрорагах.
Нематоцисты разряжаются только при
соприкосновении
с
«чужими» актиниями, не реагируя на
касания тела особей своей
колонии;
как правило, крупные особи нападают
первыми и обычно
побеждают.
Агонистическое поведение характерно
для гидробион-
тов
с территориальным распределением.
Прямые столкновения ча-
сты,
когда устанавливаются иерархические
отношения между осо-
бями.
Наряду
с прямой борьбой широко распространена
и взаимопо-
мощь.
Например, у беззубых китов и дельфинов
гибнущие особи
издают
особые сигналы, и находящиеся поблизости
здоровые жи-
вотные
спешат им на помощь, не давая тонущим
погружаться в во-
ду.
Самцы камчатского краба помогают
самкам во время линьки
освобождаться
от старого панциря. У раков-отшельников
нередок
взаимовыгодный
обмен раковинами, когда каждый из них
получает
более
подходящую по размеру. Особенно
многообразна и широко
распространена
взаимопомощь у особей в стаях и
скоплениях, а
также
при взаимодействии родительских особей
с потомством.
К
отношениям взаимопомощи относятся
также случаи выделения
особями
специфических веществ, стимулирующих
метаболизм дру-
гих
представителей своего вида или несущих
им полезную инфор-
мацию.
Например, видоспецифические вещества,
выделяемые мол-
251
люсками
Вютрка1аг1а
ц1аЪга1а,
повышают интенсивность питания
и
темп роста его молоди. Стимуляция роста
метаболитами конспе-
цифических
(своего вида) особей обнаруживается у
многих рыб-
Многие
гидробионты при ранении хищником
выделяют особые «ве-
щества
испуга», которые предупреждают об
опасности других осо-
бей
и вызывают у них реакцию бегства.
Чувствуя опасность, мол-
люск
Каьапах
тегтьз
выделяет секрет «желтой железы», что
вы-
зывает
у следующих за ним животных изменение
направления дви-
жения.
Конкуренция
и биохимическое ингибирование. Поскольку
жиз-
ненные
потребности особей одной популяции
во многом совпадают,
между
ними часто возникает конкуренция за
недостающие ресур-
сы.
В первую очередь это относится к пище
(когда ее количество
ограничено),
свету при взаимозатемнении, к субстратам,
использу-
емым
для прикрепления, размножения и др.
Например, во время
нереста
не каждая пара кеты или горбуши может
найти место,
удобное
для откладки икры. Оседающие на фукусах
личинки поли-
хеты
ЗрьгогЫз
прикрепляются друг от друга на расстоянии
1—2 мм„
что
обеспечивает достаточное пространство
для взрослых живот-
ных;
личинки, не нашедшие места, погибают.
В
какой-то мере острота внутривидовой
конкуренции снижается
в
результате дифференциации потребностей
у особей разного раз-
мера,
пола и физиологического состояния,
пространственного и вре-
менного
разграничения пользования дефицитными
ресурсами. На-
пример,
особи разного возраста обычно отличаются
по спектру пи-
тания,
держатся в разных местах и имеют
неодинаковый суточный
ритм
потребления корма. Благодаря этим
различиям конкуренция
за
пищу становится менее острой. Сглаживанию
остроты конкурен-
ции
или ее полному снятию способствуют
территориальность рас-
пределения,
иерархия взаимоотношений и регуляция
численности
популяций.
Например, моллюски Ра1е11а
сосЫеагьз
у мыса Доброй
Надежды
образуют на скалах скопления с
максимальной биомас-
сой
около 125 г/м2
и численностью до 450 экз./м2.
Когда численность
моллюсков
повышается, биомасса остается той же
за счет снижения
средней
массы особей; размер гонад и соответственно
плодовитость
животных
снижаются, когда их численность
превышает 450 экз./м2,
и
эта зависимость, играющая роль обратной
связи, контролирует
плотность
популяции, сглаживает остроту конкуренции
между осо-
бями
(ВгапсЬ, 1975). Высокая численность
моллюсков исключает
поселение
других видов пателл и обрастание
литотамний (основной
пищи
пателл) другими водорослями.
Другой
путь устранения конкуренции нередко
реализуется само-
регуляцией
численности особей на основе их
биохимического вза-
имодействия.
Как показали многочисленные исследования,
гидро-
бионты
выделяют видоспецифические метаболиты,
которые в кон-
центрациях
выше некоторых критических ингибируют
рост боль-
шей
части-особей. Рост разных особей
подавляется неодинаково,
поэтому
их размерная вариабельность возрастает.
Таким образом
обеспечивается
не только снижение суммарного прироста
биомассы
252
популяции,
но и повышение разнокачественности
особей, т. е. умень-
шение
остроты конкуренции между ними.
Например, в переуплот-
ненных
культурах хирономид отдельные личинки
обнаруживают
рекордный
рост, быстро окукливаются и превращаются
в комаров.
После
этого в процесс быстрого роста включаются
некоторые дру-
гие
личинки, окукливаются и таким образом
обеспечивается перио-
дизация
использования пространственных
возможностей, снижаю-
щая
остроту конкуренции. Достаточно в
группу мелких головасти-
ков
посадить одного крупного, как их рост
прекращается; тот же эф-
фект
наблюдается, если к мелким головастикам
приливалась вода,
где
содержались крупные особи. Сходные
результаты получены &
опытах
с карпами и другими рыбами (Константинов,
Пелипенко,.
1985).
Образование
стай и скоплений. Одно из проявлений
внутрипопу-
ляционных
отношений — объединение отдельных
особей в группы,
нахождение
в которых обеспечивает организмам те
или иные пре-
имущества.
Объединение может быть кратковременным,
например
для
совместной охоты, или охватывать
значительный период време-
ни,
как это наблюдается у многих бентосных
форм. В свою очередь*
сильно
различается и степень интеграции
особей в группах. В свя-
зи
с этим классификация форм объединения
до настоящего време-
ни
разработана слабо и многие используемые
в этой области поня-
тия
недостаточно точны. Объединения
пелагических животных обыч-
но
принято называть стаями, бентосных —
скоплениями, агрега-
циями.
Ярким
примером объединений бентосных животных
могут слу-
жить
сложно организованные скопления
морских ежей, офиур, го-
лотурий.
Морские ежи ЕсЫпагасктиз,
МеИИа, БепЛгавШ,
питаю-
щиеся
сестоном, лежат друг на друге в несколько
слоев, что прежде
всего
улучшает им условия питания. Возникающие
вторичные токи
воды
усиливают поступление сестона,
повышается общая ловчая
поверхность,
так как особи верхних слоев, лежа на
грунте, могут-
фильтровать
воду всей поверхностью тела. Особи
нижних слоев не-
поедаются
морскими звездами, молодь со слабыми
иглами нахо-
дится
под защитой взрослых ежей. В скоплениях
создается высокая
концентрация
половых продуктов, что повышает
эффективность
размножения
животных. В плотных скоплениях морских
лилий и
офиур
лучи, расправленные в потоке, способствуют
улавливанию
пищевой
взвеси и стабилизации силы течения.
Сходная картина
описана
для ряда голотурий.
Своеобразные
шарообразные скопления образуют крабы
Ма}а
вдитайо,
в которых слинявшие самцы располагаются
по периферии
большой
кучи (до 1 м в диаметре), а неслинявшие
самцы и самки —
в
середине, где они линяют под защитой
внешнего слоя животных-
В
кучах крабы так переплетаются ногами,
что осьминоги не в си-
лах
оторвать отдельных животных, захватывая
иногда лишь те
особи,
которые недостаточно крепко «вмонтированы»
в общую
массу.
Скопление устриц, в частности, полезно
для их размножения
и
выживания молоди, которая, оседая из
толщи воды, прикрепля-
253
ется
к раковинам взрослых особей. То же
наблюдается у дрейссены.
Даже
нанесение водной вытяжки устричной
ткани на поверхность
субстрата
стимулирует оседание на него молоди;
аналогичное яв-
ление
отмечено для молоди усоногих. Поскольку
усоногие имеют
внутреннее
оплодотворение, часто облигатно
перекрестное, скопле-
ние
особей обеспечивает их размножение.
Так как для рачков ха-
рактерны
свободноплавающие планктотрофные
личинки, излишнее
рассредоточение
особей предупреждается стайным
оседанием на
раковины
конспецифических особей. Предпочтительное
оседание
личинок
на субстраты, уже занятые представителями
своего вида,
■отмечено
у моллюсков СоИвеИа,
морских блюдечек, голотурий, ас-
цидий,
червей.
Формирование
стай очень характерно для пелагических
живот-
ных.
Большие скопления — «флотилии» —
образуют на поверхности
морей
водомерки На1оЪа1ез
гоЪизЫз.
Приближение врагов вызыва-
ет
у них резкое возрастание скорости
движения, частоты поворотов,
столкновения
особей и мелькание бликов света, что
дезориентирует
хищников,
прицельно схватывающих жертву. По
эксперименталь-
ным
данным, логарифм числа нападений сардин
в расчете на одну
водомерку
был обратно пропорционален числу
насекомых во флоти-
лии
(Ро8{ег, ТгеЬегпе, 1981). Огромные стаи
численностью несколь-
ко
миллионов особей и площадью до 20—30 м2
образуют среди ко-
ралловых
рифов веслоногие АсагНа,
ОИкопа
и Сеп&орадез.
Веду-
щую
роль в образовании их стай играет
защита от выедания; ночью
стаи
рассредоточиваются, утром образуются
вновь. В рое наупли-
усов
ЕШтшв
тойезЫз,
располагающемся у поверхности воды,
за
счет
движения животных создаются общие
токи воды, усиливающие
индивидуальные
(улучшение условий питания и дыхания).
Стаи из
сотен
и тысяч самцов образуют крабы-плавунцы
Ро1уЫив
кеп$1ош
и
СкагуЬсИ8
ейхюапйзь.
В Антарктике обнаружены скопления
криля
протяженностью
несколько километров и толщиной
несколько сот
метров;
общая биомасса рачков по акустическим
данным достигала
10
млн. т, концентрация— 10 тыс. экз./м3
(5Ьи1епЬег§ег, 1983).
У
пелагических мизид стаи формируются
отдельно для молодых
и
половозрелых особей в соответствии с
различием скорости их дви-
жения.
Параллельная плавательная ориентация
достигается зри-
тельно
и путем тигмотаксиса — восприятия
токов воды, создава-
емых
при плавании, дыхании и питании отдельных
особей. Стай-
ность
взрослых мизид создает условия,
благоприятные для их раз-
множения:
самки способны к оплодотворению (каждые
10 дней) в
течение
всего 5 мин и потому должны держаться
вблизи самцов.
На
восприимчивость самок к оплодотворению
влияет плотность
скоплений:
с ее уменьшением процент беременных
самок в стаях
снижается.
У
многих кальмаров Отта1о81геркв8
р1егори.в
стаи
могут вклю-
чать
до 1000 особей сходного размера; во время
охоты стаи распа-
даются,
затем снова восстанавливаются.
Многие
животные образуют в толще воды брачные
рои, кото-
рые
помогают особям разного пола находить
друг друга. Так, с на-
254
ступлением
темноты всплывают в толщу воды и
формируют брач-
ные
рои многие бентосные раки. Грандиозное
роение наблюдается
у
донных червей Ейтсе,
когда они огромными массами всплывают
для
размножения к поверхности моря. Такое
же роение происходит
у
червей Ыегегз
1оп$г18з1та.
Особенно
характерно стаеобразование для рыб.
Благодаря внут-
ристайной
сигнализации стайные рыбы имеют
определенные пре-
имущества
перед одиночными в количестве информации
об окружа-
ющем
мире. Нахождение в стае помогает
животным обнаруживать
врагов
и защищаться от их нападения, так как
животные в груп-
пах,
состоящих из особей с разными навыками,
в целях самосохра-
нения
используют не только свой, но и
коллективный опыт.
Пребывание
в стае помогает рыбам эффективнее
спасаться от
преследования
хищников. Так, из стайки молоди сайды
первая рыб-
ка
оказалась пойманной треской через 2
мин 15 с, тогда как оди-
ночные
сайды становились жертвой через 26 с.
Это объясняется
тем,
что хищник, нападая на стаю, не преследует
какую-либо одну
рыбку,
а, погнавшись сначала за одной и упустив
ее, устремляется
за
другой, за третьей, все время имея дело
с новыми, не утомленны-
ми
погоней особями. Подобное явление
наблюдается и при атаке
стай
рыб кальмарами, каракатицами.
Оборонительный
маневр стаи обычно заключается в том,
что-
рыбки
быстро и согласованно разбегаются в
стороны перед хищни-
ком,
огибая его, и снова сплачиваются. Это
дезориентирует хищ-
ника,
исключает прицельные броски и затрудняет
охоту. Сильное
дезориентирующее
действие оказывает на хищника
взрывообраз-
ное
рассеивание стаи в момент атаки на
нее.
В
темноте рыбы не могут согласованно
маневрировать и вместе
с
тем легче обнаруживаются хищниками с
помощью обоняния. По-
этому
вечером стаи рассредоточиваются, а
утром формируются за-
ново.
Новое формирование происходит за счет
объединения особей,
ранее
входивших в состав разных стай. Благодаря
этому в новук>
стаю
входят особи с разным опытом, суммарный
фонд условных
рефлексов
возрастает и становится достоянием
всех рыб стаи.
Нередко
стаи рыб и других гидробионтов образуются
не для
обороны,
а нападения. Например, солнечники
объединенными уси-
лиями
атакуют крупных инфузорий, косатки
собираются в стаи для
охоты
на крупных китов. Желтохвосты, нападая
на стаи ставриды,,
идущие
вдоль береговой линии, отсекают группы
в несколько сот
рыб,
направляют их на мелководье, где
окружают и уничтожают;
преследуя
рыб, обитающих в прибрежных рифах,
желтохвосты груп-
пой
из 10—15 экземпляров отсекают часть
стаи, отгоняют ее в от-
крытые
воды, где без труда овладевают своей
жертвой (ЗсЬгшМ,
81гап(1,
1982).
Продукция
органического вещества
и
трансформация энергии
Каждая
популяция синтезирует органическое
вещество, которое
накапливается
в особях, содержится в выделяемых ими
метаболи-
тах,
в выметываемых половых продуктах,
отчуждаемых структурах,
используется
паразитами. Темп продуцирования
органического ве-
щества
популяциями определяется как их
биологическими особен-
ностями,
так и всем комплексом факторов внешней
среды. Энерге-
тическое
и материальное обеспечение продуцирования
осуществля-
ется
в разных формах автотрофного и
гетеротрофного питания.
Поступающая
в популяцию энергия, трансформируясь
с той или
иной
эффективностью, образует поток, величина
которого и его от-
дельные
направления в обобщенной форме
выражаются уравнени-
ем
энергобаланса.
Темп
и эффективность продуцирования. Темп
продуцирования
популяциями
органического вещества характеризуется
скоростью и
интенсивностью
процесса. Мерой скорости продуцирования
служит
количество
органического вещества, образуемое
популяцией в еди-
ницу
времени. Интеграл скорости продуцирования
выражает вели-
чину
продукции,
т. е. количество органического вещества,
создава-
емого
за весь рассматриваемый отрезок
времени. Мерой интенсив-
ности
продуцирования служит удельная
продукция
— количество
-органического
вещества, образуемого популяцией в
единицу време-
ни
на единицу массы. Нередко величину и
скорость продукции рас-
считывают
не для всей популяции, а лишь для той
ее части, которая
занимает
единицу пространства (например, на 1
м2
или 1 м3).
Та
часть органического вещества, которая
аккумулируется в
«особях,
называется биологической
продукцией.
Ее величину выра-
жают
биомассой, вновь образуемой всеми
особями за рассматрива-
емый
период времени вне зависимости от
того, какую часть этого
периода
они существовали в популяции. Величина
биологической
продукции,
отнесенная ко времени, отражает скорость
продуциро-
вания.
Показателем интенсивности продуцирования
служит Р/В-ко-
.эффициент
— отношение прироста биомассы
(продукции) за иссле-
дуемый
промежуток времени к ее средней величине
за тот же срок.
Следовательно,
Р/В-коэффициент характеризует удельную
биопро-
дукцию
популяций.
Продукционные
показатели популяций в первую очередь
зави-
сят
от их структуры и видовых особенностей
организмов. Как пра-
вило,
с увеличением продолжительности
жизненного цикла гидро-
бионтов
интенсивность продуцирования ими
органического вещест-
ва
снижается. Например, у бокоплавов
Саттагив
1осив1а, О. 1а-
яивМв
и АсапШо^аттагив
дгеычпдМ,
живущих соответственно
менее
1 года, около 2 и около 10 лет, суточный
Р/В-коэффициент
равен
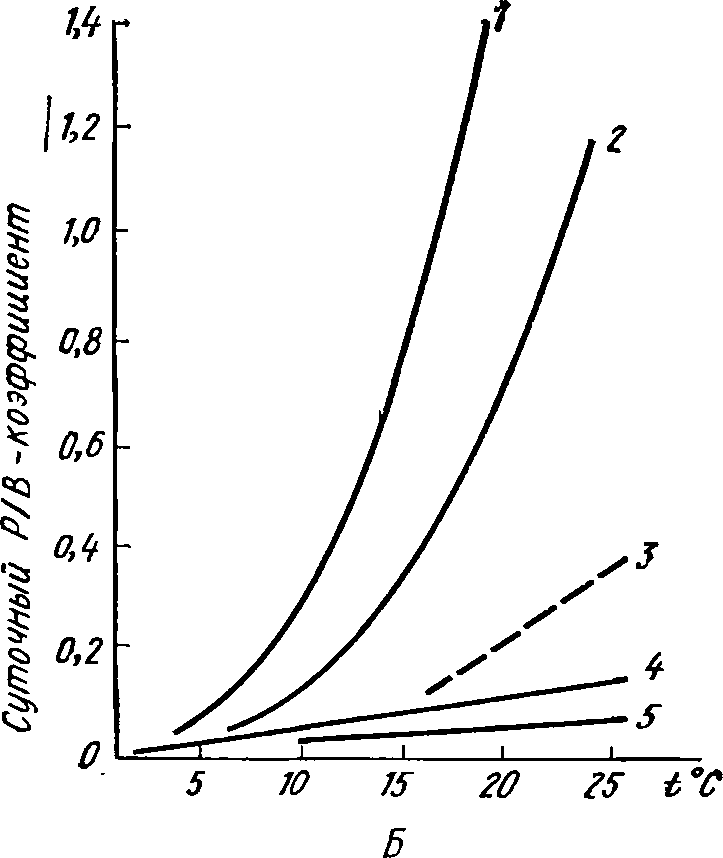
0,048; 0,0055 и 0,0004. На рис.43 хорошо видно,
как закономер-
но
уменьшается Р/В-коэффициент у ряда
других гидробионтов по
мере
перехода от короткоживущих форм к
особям, имеющим боль-
шую
продолжительность жизни. Для многих
представителей мор-
256
ского
макробентоса рассматриваемая зависимость
аппроксимирует-
ся
уравнением (КоЬейбОП, 1979):
1дР/й=0,66—0,721д^, где
I
—
продолжительность
жизни (годы).
Органическое
вещество интенсивнее продуцируется
в популя-
циях,
образованных мелкими организмами. У
бактерий суточный
Р/й-коэффициент
нередко выражается единицами, несколько
ниже
он
у бесцветных жгутиковых, у инфузорий
лежит в пределах
0,5—3,
у коловраток не превышает нескольких
десятых долей. Еще
меньше
он у низших ракообразных (0,02—0,45), высших
раков
(0,0014—0,05),
моллюсков (0,0001—0,003), иглокожих
(0,0007—
0,022)
и рыб (до 0,08). Приведенные величины
вычислены приме-
нительно
ко всему жизненному циклу организмов.
На ранних ста-
диях
развития гидробионтов интенсивность
продуцирования ими
органического
вещества выше, на поздних — ниже средних
величин.
Иногда
необычайно высокий темп продуцирования
наблюдается у
сравнительно
крупных животных. Например, сальпы в
благоприят-
ных
условиях увеличивают свою массу на
10% в час, за сутки
часто
удваивают ее (Кашкина, 1975).
Поскольку
темп роста особей находится в обратной
зависимости
от
массы, продукция популяции будет тем
выше, чем больше в ее
составе
окажется молоди. В значительной мере
скорость продуци-
рования
снижается в моменты размножения,
особенно если в
это
время особи частично или полностью
прекращают пи-
тание.
Скорость
продуцирования возрастает с повышением
экологиче-
ской
обеспеченности роста особей, с улучшением
условий сущест-
вования
гидробионтов (в первую очередь
трофических, температур-
ных
и кислородных). Например, в зимнее время
для многих бес-
позвоночных
Р/В-коэффициент приближается к 0, весной
и осенью
он
заметно ниже, чем летом. На рис. 43 видно,
насколько различно
изменяется
величина Р/й-коэффициента у гидробионтов
с повыше-
нием
температуры. Обычно эта зависимость
имеет линейный или
степенной
характер. Например, с повышением
температуры от 8 до
22°
С Р/й-коэффициент мелких инфузорий
возрастает в 10 раз, сред-
них
— в 8 и крупных — в 6 раз (Заика, 1983).
Эффективность
продуцирования органического вещества
опре-
деляется
отношением его количества, синтезируемого
популяцией,
к
тому, которое используется ею. Количество
использованного ор-
ганического
вещества можно выразить рационом
популяции или
его
частью, ассимилируемой особями.
Получаемые показатели К\
и
К'2
аналогичны трофическим коэффициентам
К\
и К2,
но выше
их
по абсолютному значению и имеют другой
экологический смысл.
Конкретные
значения К\
и К'2
зависят от видовых особенностей
организмов,
структуры популяций (в первую очередь
возрастной
и
размерной) и внешних условий (в первую
очередь трофических,
температурных
и респираторных).
Одним
из показателей эффективности
продуцирования служит
соотношение
между ассимиляцией и энерготратами,
выраженное в
эквивалентных
величинах. Оно может равняться нулю,
когда вся
а—2062
257
I
53
53
&
I
I
■=0 10 и) 0,1 от ■51 1 0Д11 0,0001 |
Д |
|
|
|
|
Л |
Всю □ |
|
|
д 7 □ 2 о 3 |
|
|
ей да |
3 т |
• * ■ 5 |
|
|
^°с§ □ |
1о- ■ |
|
|
|
|
и -Г* |
0,1
1,0 Ю 100 1000
время,
сутки
Рис.
43. Удельная биопродукция гидробионтов
(по
Заике, 1983). А—суточный
Р/В-коэффи-
циент
у некоторых гидробионтов с разной
про-
должительностью
жизни:
/
— черви, 2
— ракообразные, 3
— моллюски, 4
— игло-
кожие,
5 — рыбы, 6
— полоса наиболее обычных зна-
чений
для инфузорий;
Б
— суточный Р/В-коэффициент у
некоторых
гидробионтов
при разной температуре:
1
— инфузории, 2
— коловратки, 3
—
гарпактициды,
4
— нематоды, 5 — олигохеты
ассимилированная
пища
расходуется
только на
поддержание
биомассы
(поддерживающий
раци-
он).
Такой случай наблю-
дается,
когда животные
находятся
на голодном
рационе
или прекращают
рост
под влиянием каких-
либо
неблагоприятных
абиотических
условий.
С
улучшением условий
отношение
продукции к
энерготратам
возрастает,
повышается
оно и тогда,
когда
в популяциях уве-
личивается
относительная
численность
младших воз-
растных
групп.
Энергобаланс
популя-
ций.
Энергобаланс каждой
популяции
отражает ее
специфические
свойства, а
также
находится под влия-
нием
внешних условий.
Например,
в водоеме Кра-
пина
(Румыния) средняя
биомасса
беззубки А.
р1-
зстаИз
составляла 28,9
г/м2
или 57 Дж/м2,
интен-
сивность
дыхания — 94
Дж/м2
в год, фиксирован-
ная
в популяции энергия
достигала
10% от ассими-
лированной.
В том же во-
доеме
популяция перлови-
цы
V.
рШогит
имела био-
массу
15,9 г/см2
или
47
Дж/м2,
в процессе ды-
хания
рассеивалось 121
Дж/м2
в год из 135 Дж/м2
ассимилируемых,
т. е. око-
ло
6% усваиваемой энер-
гии
аккумулировалось и
94%
рассеивалось.
У
популяции морских
ежей
51гопцИосеп1го1и8
ЛгоеЬасЫепвгв,
обитающих
у
побережья Новой Шот-
258