

20
1
*'
4 12
20 28 36 И
О
Б
Рис.
37. Кривые роста животных. А—самки
БарНЫа
ри1ех
(из Заики, 1972):
/
— соматический рост. 2
— генеративный рост. 3
— сумма массы тела и всех отложенных
к
дгкиому времени яиц;
Б
— гребневика Вегоё
&гасШз
и 8ацШа
ЫзрШа:
I
— для-одной особи В.
вгасШз, 2
— средняя для трех особей В.
%гасШз, 3
— для сагитты
ского
вещества, сухой массы, зольных элементов).
По продолжи-
тельности
различают бесконечный,
или ассимптотический,
рост, для-
щийся
всю жизнь, и конечный,
завершающийся по достижении ор-
ганизмом
какого-то возраста (массы). Если в
онтогенезе размер
особи
увеличивается с сохранением
геометрического подобия, го-
ворят
об изометрическом
росте в отличие от
анизометрического
(аллометрического),
когда пропорции тела с возрастом
меняются.
При
строго изометрическом росте объем (№)
возрастает пропорцио-
нально
кубу длины (Ь)
или иного линейного показателя, т. е.
№=
а-Ь3,
где а
— коэффициент пропорциональности. В
случае анизо-
метрического
роста уравнение принимает вид УР=а-Ьъ,
где ЬФ3.
Если
ширина и (или) высота возрастают
медленнее, чем длина,
6<3,
в обратном случае 6>3.
Рост,
при котором изменяется соотношение
массы отдельных
тканей,
называют гетерогенным,
в обратном случае — гомогенным.
При
периодическом
росте наблюдается та или иная форма
преры-
вистости
процесса, связанная с изменением
внешних условий или
физиологического
состояния организмов. Например, у
многих дву-
створчатых
моллюсков выделены суточные, приливные
и годовые
циклы
роста, хорошо прослеживающиеся по
частоте линий нара-
стания
раковины, в том числе их сгущения
(замедление роста) в
момент
полового созревания (Ап1оше, 1980).
Постоянный
рост ха-
рактеризуется
непрерывностью (вплоть до отмирания),
хотя его
интенсивность
сильно зависит от условий, стадий
онтогенеза и обыч-
но
замедляется после наступления
половозрелости.
222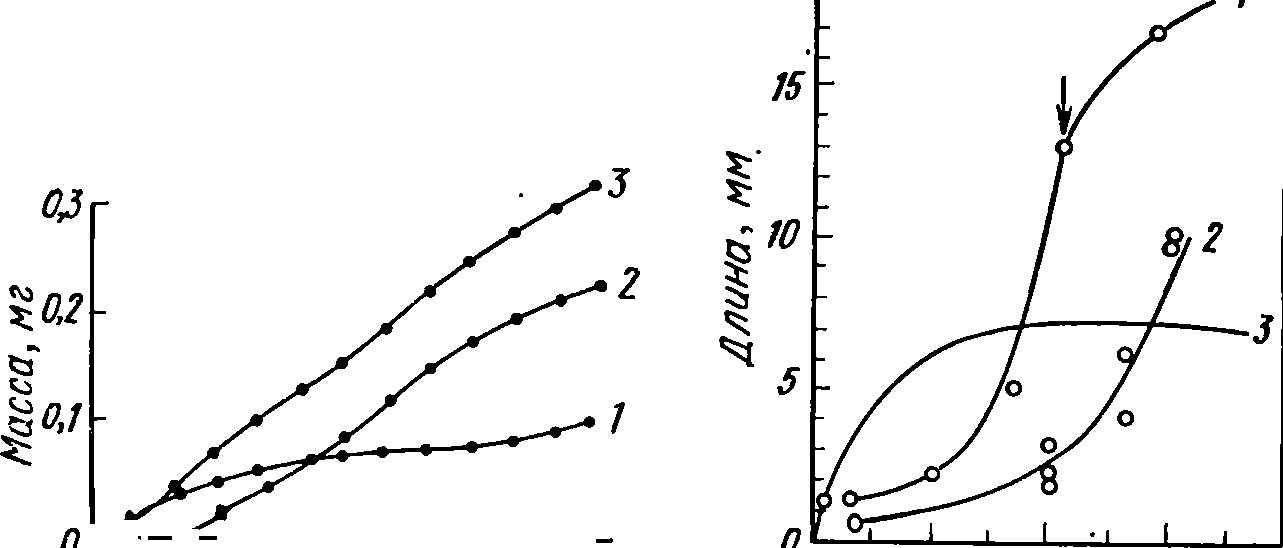
п*3-
* ' 1 1 1 1—10 20 30 Ы) Возраст, сутки
Возраст, сутки а
5. М&рьйа
Для
математического описания роста животных
на протяжении
всего
онтогенеза часто используют уравнение,
основанное на раз-
работках
А. Пюттера, Л. Берталанфи, К. Тейлора и
некоторых дру-
гих
авторов. Оно исходит из положения, что
рост — это разность
между
ана- и катаболизмом:
—=Л1Г
а(
где Л — «константа анаболизма», К — «константа катаболизма»,
масса организма, т и п — показатели степени. Принимая, что п= 1, и интегрируя (1), получаем •
1
1Г<=[1Гсо(1-е<1-т>*<)]1-т. (2)
где — масса во время 1\ №«> — конечная масса организма, рас- сматриваемая как (Л :К)1~т •
В. Е. Заика (1972) показал, что параметрам уравнения (1) мож- но придать определенный биологический смысл. А№т можно рас- сматривать как величину ассимиляции при условии, что она всегда равна одной и той же части рациона и степенной зависимости ра- циона от массы тела. К№п представляет собой величину энерготрат (дыхания) организма с массой №. Если т = п, рост будет парабо- лическим, при т<п получается 5-образная кривая (см. рис. 37), а при т=п= 1 рост экспоненциальный.
В случае изометричности линейный рост исходя из (1) описы- вается уравнением
(3)
где и — конечная длина, достигаемая организмом, 10 — возраст, в котором 0. Например, для морского ежа 8ргоп§ИосеШгоШз т- 1егте(Ииз уравнение (3), согласно Э. Брегману (11971), имеет сле- дующие параметры:
Ь(= 69,2 [ 1 - 6-0,881(1-^219)^
где — диаметр панциря (мм), I — возраст (годы). Поскольку масса и длина ежа связаны уравнением №=0,527 Ь2-д37у для описа- ния роста сырой массы ежа получаем уравнение
УК/= 134000 [1 — е -0,331«-0,219)]2,9375
где длина выражена в миллиметрах, масса — в миллиграммах.
Кривая роста 5-образной формы означает, что абсолютные при- росты массы в единицу времени сначала невелики, а затем стано- вятся все большими, достигают максимума, после чего начинают снижаться, асимптотически приближаясь к 0. Иначе идет измене- ние относительных приростов, которые с возрастом обычно снижа- ются. Иногда относительные приросты в онтогенезе закономерно возрастают, как, например, это описано для некоторых медуз.
223
Построение
кривой роста по эмпирическим данным,
полученным
на
группе особей, может дать неверное
представление об индивиду-
альном
росте, как это видно на рис. 37. У личинок
хирономид ха-
рактер
роста отдельных особей вплоть до
окукливания прямолине-
ен,
а по средним данным для всей группы —
5-образный вследствие
выхода
из нее наиболее крупных особей в
результате окукливания
(Константинов,
1956).
На
протяжении всего онтогенеза рост
нельзя достаточно точно
выразить
единым уравнением. Лучшие результаты
дает его описа-
ние
применительно к отдельным отрезкам
онтогенеза. Как пока-
зал
И. И. Шмальгаузен (1935), в этом случае
рост хорошо аппрок-
симируется
уравнением ^2=^11к,
где и 1^2
— масса организма
в
начале и конце рассматриваемого
периода, I
— его длительность,
к
— «константа роста», характерная для
организмов каждого вида.
В
качестве показателя темпа роста И. И.
Шмальгаузен (1927)
предложил
использовать удельную
скорость роста (С\у),
вычисляе-
мую
по формуле
, (4)
‘2
— Н
где и №2 — масса организма в начале (/]) и конце (^2) рассмат- риваемого периода. В подавляющем большинстве случаев удельная скорость роста снижается с увеличением массы животных (Л<1).
С возрастом пропорции тела гидробионтов редко остаются не- изменными, равно как и соотношение отдельных органов, тканей и их биохимический состав. У ряда водных беспозвоночных и рыб выявлена скачкообразность в развитии аллометрии и гетерогении. Как показал В. В. Васнецов (1947), у рыб смена периодов изомет- рического роста аллометрическими процессами приводит к синхрон- ной перестройке сразу целого комплекса структур.
Приспособительность роста. Аллометрия и гетерогенность роста имеют приспособительное значение и довольно заметно влияют на выживаемость особей и смертность популяций. Например, преиму- щественный рост в высоту, обусловливающий высокотелость рыб, резко снижает выедаемость их хищниками. Аллометрия роста бес- позвоночных, особенно сопровождающаяся образованием всевоз- можных шипов, выростов, колючек, играет огромную защитную роль и сильнее выражена в условиях повышенного пресса хищни- ков. По этой причине вооруженность особей в популяциях тропиков, где относительное количество хищников очень велико, выше, чем в умеренных и приполярных зонах.
Приспособительное значение гетерогенности роста хорошо про- слеживается на примере жиронакопления. В тропиках, где условия питания на протяжении года сходны, жирность особей в популяции колеблется в небольших пределах. В высоких широтах в благопри- ятные для питания периоды рост особей сопровождается образова- нием большого количества жира, последующее использование ко- торого во время перерыва в питании повышает их выживаемость. Например, среднее количество жира у планктонных организмов от
224
112°
с. ш. до 8° ю. ш. равно 8,7%, а севернее 25°
и южнее 26° повы-
шается
соответственно до 14,5 и 19,5%.
Сходная
картина прослеживается у большинства
рыб. У сигов
в
северной части их ареала сезонный ритм
жиронакопления выра-
жен
слабее, чем в южной, где имеет место
перерыв или значитель-
ное
снижение интенсивности питания в
жаркое время года (Пирож-
ников,
1955).
Анализируя
биологические закономерности роста
рыб, Г. В. Ни-
кольский
(1974) отмечает, что быстрый рост
способствует более
раннему
наступлению половозрелости и меньшему
выеданию хищ-
никами.
Более быстрый, чем в предыдущий период
жизни, темп
увеличения
массы тела у половозрелых особей
обеспечивает более
интенсивное
наращивание плодовитости популяции.
До наступле-
ния
половозрелости основные энергетические
ресурсы, поступаю-
щие
в организм, расходуются на белковый
рост и увеличение дли-
ны
особей; после — на наращивание массы
тела и накопление ре-
зервных
веществ. Отмеченные закономерности
характерны и для
многих
беспозвоночных.
Конечный
размер гидробионтов видоспецифичен
и, как все ви-
довые
признаки, адаптивен. Малые размеры
облегчают отыскание
убежищ,
позволяют существовать за счет
ограниченных кормовых
ресурсов,
быстрее достигать репродуктивной
зрелости, давать боль-
шее
число поколений и маневреннее
использовать колеблющиеся
условия
среды. Крупные размеры снижают
выедаемость особей,
позволяют
относительно большему их числу достигать
репродуктив-
ного
возраста, давать многочисленное
потомство, уменьшать зави-
симость
воспроизводства от кратковременных
неблагоприятных из-
менений
среды.
Влияние
различных факторов. Генетическая
программа роста,
различная
даже у особей одного вида, в той или
иной мере реализу-
ется
в пределах, обусловливаемых внешними
условиями. К основ-
ным
абиотическим факторам, влияющим на
реализацию потенций
роста,
относятся температура, свет, газовый
режим воды, раство-
ренные
в ней соли, гидродинамика. Из биотических
факторов наи-
большее
значение имеет трофический, а также
аллелопатия, агони-
стические
и иерархические взаимоотношения.
С
повышением температур до некоторых
оптимальных значений
темп
роста ускоряется, после чего снижается.
Например, суточные
приросты
раковины у моллюска Мегсепапа
тегсепаг1а
при 7,5 и
32°С
равны 30 мкм, а в середине этого интервала
(20°С) достигают
200
мкм, причем восходящая и нисходящая
ветви кривой зеркально
подобны
друг другу. Личинки хирономид СЫгопотиз
йогзаНз
при
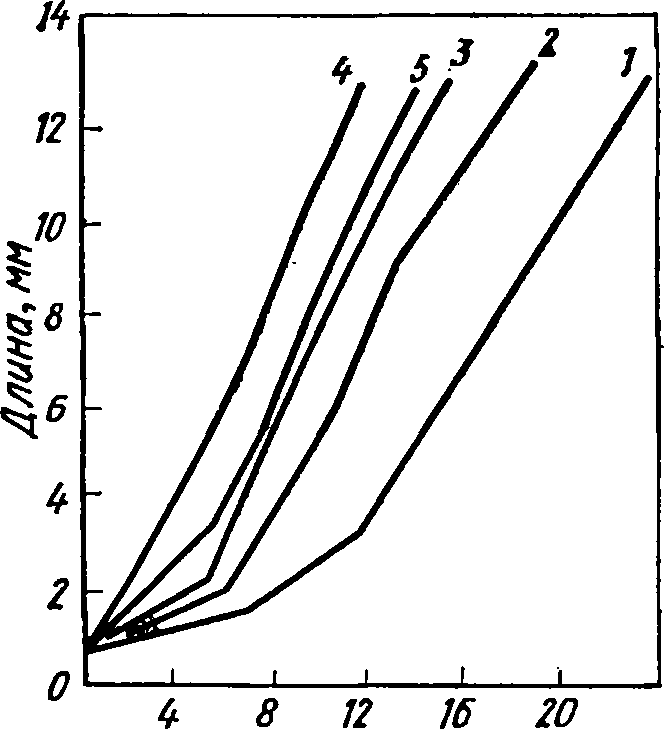
15,
20, 25 и 30° С достигают массы 12 мг
соответственно за 17, 13, 9
и
13 дней (рис. 38). Скорость роста креветок
Сгащоп
зер1епзрто-
за
в водах Род Айленд (США) обнаруживает
хорошо выраженную
линейную
зависимость от температуры.
Одновременно
с ускорением роста под влиянием высоких
темпе-
ратур
наблюдается уменьшение конечных
размеров животных.
В
приведенном примере с СН.
йогзаНз
конечная масса личинок при
8—2062
225
115,
20, 25 и 30°С равнялась со-
ответственно
11,1; 9,4; 7,4 и
мг,
т. е. практически сни-
жалась
вдвое. Инфузории ту-
фельки,
полученные от одной
особи,
при 18 и 28°С имели дли-
ну
соответственно 187 и
230
мкм. Масса планарий воз-
растала
с 25 до 35 и 53 мг, ког-
да
температура снижалась с 18
до
12 и 7РС.
Следует отметить,
что
в природных условиях наи-
лучший
рост животных может
наблюдаться
при иных темпе-
ратурах,
чем в эксперименте,
вследствие
влияния многих
факторов
одновременно. Полу-
чены
данные о том, что в коле-
бательном
терморежиме гидро-
бионты
растут лучше, чем в
стабильном
(см. гл. II).
Как
правило, с изменением
температуры
меняется алломет-
рия
роста. Например, у личи-
нок
морских ежей 8(гопдИо-
сеп1го1из
с повышением температуры от 11,4 до 15,9;
20,4 и 23,7°С
длина
анальных придатков возрастала на 30,
36 и 27%, у 5ркаеге-
сктив—
на 178, 324 и 387%. Сходная картина изменения
относи-
тельной
длины различных структур наблюдается
у многих простей-
ших
и других животных.
Существенно
влияет температура на биохимический
состав тела.
Например,
у карпов, выращиваемых в водоемах-охладителях,
усили-
вается
синтез липидов, и их содержание в тканях
заметно повыша-
ется
(Романенко, 1983). В природных условиях
для рыб и многих
беспозвоночных
характерно жиронакопление, связанное
с зимовкой
или
иными периодами жизни, когда питание
выключается и энерге-
тика
обеспечивается только за счет внутренних
резервов.
Свет,
основной фактор роста в жизни растений,
далеко не без-
различен
и для животных. Например, рост и линька
крабов Сесаг-
стиз
ШегаИз
подавляется при круглосуточном
освещении, многие
планктонты
держатся у поверхности воды, нуждаясь
в свете для
синтеза
витаминов. В эксперименте молодь нерки
и чавычи в тем-
ноте
росла хуже, чем при круглосуточном или
12-часовом освеще-
нии,
то же характерно для ушастого окуня.
Канальные сомики, ак-
тивные
преимущественно ночью, лучше всего
растут в полной тем-
ноте.
Сильнейшим
депрессором роста для животных может
быть недо-
статок,
а для растений — избыток кислорода. С
повышением его
концентрации
до 3—4 мг/л включается фотодыхание
растений, уси-
Возраст,
сутки
Рис.
38. Линейный рост личинок СМ-
гопотив
йог$аИ$
при разной темпера-
туре:
Г
— 10°, 2 — 15°, 3
— 20’, 4
— 25°, 5 — 30° С
226
ливающееся
с ростом аэрированности среды, и
продуктивность
фотосинтеза
резко снижается (Лархер, 11978). Высокие
концентра-
ции
СОг и НС03-,
благоприятствующие росту растений, в
некото-
ром
диапазоне имеют то же значение для
животных. Как показал
В.
Д. Романенко с соавторами (1980), у рыб
скорость реакций кар-
боксилирования
с использованием СОг из окружающей
среды воз-
растает
по мере повышения концентрации
углекислоты до некото-
рых
оптимальных значений, и соответственно
усиливается биосин-
тез
различных соединений за счет гетеротрофной
фиксации углеро-
да.
У карпов 14С
из №Н14СОз
наиболее интенсивно включался в
состав
тканей печени, когда уровень С02
достигал 8—18 мг/л, т. е.
в
несколько раз превышал ее нормальное
содержание в воде. О вли-
янии
на рост солености, концентрации
водородных ионов, Н25
и
других
гидрохимических факторов уже говорилось
выше (см. гл.1).
Рост
донных растений и животных в значительной
мере зависит
от
особенностей грунта. Например, прирост
моллюска Муа
агепагса
на
иле, песке и гравии составлял
соответственно 2,45; 1,77 и 0,93 мг
сухой
массы в день; раковины моллюсков,
выросших на гравии,
имеют
более округлую форму и наибольшую
относительную массу
{Ые\уе11,
Шйи,
1982).
В
водоемах умеренной зоны четко проявляются
сезонные особен-
ности
роста. Летом гидробионты, усиленно
питаясь, растут наибо-
лее
интенсивно. С приближением осени резко
возрастает накопле-
ние
липидов, используемых в период зимнего
голодания. Осенью
темп
роста у гидробионтов резко снижается,
а зимой у большинства
прекращается
и постепенно возобновляется с
наступлением весны.
Огромно
влияние на рост трофических условий:
количества пи-
щи,
ее концентрации, доступности,
калорийности, биохимического
состава.
В связи с пищевыми условиями меняется
не только темп-
роста
гидробионтов, но и состав их тела.
Например, выращивание
рыб
на углеводистой пище сопровождается
их ожирением, резким
уменьшением
содержания белка. Характерная особенность
беспоз-
воночных
и рыб — высокое использование белков
на энерготраты.
Поэтому
снижение их доли в пище резко меняет
метаболизм гидро-
бионтов,
трансформирует процессы биосинтеза
и соответственно их
результат
— биохимический состав тканей. Рост
резко замедляется
или
вовсе прекращается, когда пища
недостаточно агрегирована, и
ее
добывание энергетически малоэффективно.
То же самое наблю-
дается
в случае малой доступности кормовых
объектов из-за осо-
бенностей
их строения или поведения. Интересны
данные о том, что
в
среде с небольшим количеством пищи
рост некоторых гидробион-
тов
достигает оптимального уровня при
более низких температурах
(Суханов,
1979).
Разнообразное
влияние на рост оказывают особенности
взаимо-
действия
особей. Как уже говорилось, иерархия
сопровождается
неравенством
ресурсного обеспечения особей разного
ранга и со-
ответственно
различиями в темпе их роста. Подавление
роста обыч-
но
у особей, часто подвергающихся агрессии
при разных формах
агонистического
поведения (отнятие убежища, симбионтов,
кормо-
**
227
вых
участков и др.). Очень сильно влияют на
рост неспецифические
и
специфические экзометаболиты, выделяемые
особями своего вида
и
другими организмами (см. гл. 10).
Развитие
В
экологическом плане развитие гидробионтов
в наибольшей
степени
характеризуется особенностями смены
структур и их функ-
ций
в онтогенезе, влиянием различных
факторов на скорость рас-
сматриваемого
процесса и адаптациями к выживанию
организмов
на
разных стадиях. Дефинитивное состояние
достигается путем та-
кой
последовательности сменяющих друг
друга промежуточных
форм,
которая в наибольшей мере разрешает
проблему ресурсного
обеспечения
развивающихся особей. При этом темп
развития оп-
ределяется
двумя основными тенденциями. С одной
стороны, он
максимизируется,
чтобы быстрее достичь требуемого
этапа, с дру-
гой—
может замедляться или временно
приостанавливаться в ин-
тересах
синхронизации ресурсных потребностей
и возможностей
среды.
Например, ускорение эмбрионального
развития и появление
личинок
растительноядных животных ранней
весной до того, как
в
воде станет много фитопланктона,
сопровождалось бы массовым
отмиранием
особей. Обеспечение их максимальной
выживаемости
и
по возможности полной реализации
репродуктивного потенциа-
ла—
стратегия, которой подчиняются интересы
более частного ха-
рактера.
Формы.
У водных животных, по-видимому, чаще,
чем у назем-
ных,
встречается непрямое развитие.
Какое-либо точное сравнение
тут
невозможно, поскольку в ряде случаев
трудно решить, протекает
онтогенез
организма с метаморфозом или без него.
Как показано
на
рыбах В. В. Васнецовым (1953), всякое
развитие скачкообразно
и
сопровождается синхронными перестройками
формы и функций
сразу
многих систем. Например, увеличение
длины воблы от 5 до
мм
одновременно влечет за собой изменение
формы тела, рото-
вого
аппарата, кишечника, нервной системы.
Перестройки, связан-
ные
с переходом на следующий этап у животных
с прямым разви-
тием,
часто не менее существенны, чем
наблюдаемые при метамор-
фозе.
Наиболее
резко метаморфоз выражен у гидробионтов,
развива-
ющихся
в разных средах. Например, у малоподвижных
или непод-
вижных
донных организмов пелагические личинки
приспособлены
к
иной среде и потому резко отличаются
от родительских форм. Еще
резче
выражен метаморфоз у гетеротопов,
которые в имагинальной
стадии
обитают в воздушной среде (многие
насекомые и некото-
рые
другие животные). Экологическая выгода
метаморфоза усили-
вается1
в тех случаях, когда в водоеме на
короткое время появля-
ются
источники питания, которые из-за их
нестабильности не могут
использовать
взрослые формы (например, вспышки
фитопланкто-
на
в арктических водах). Сходным образом
проявляется выгода
метаморфоза,
когда взрослые формы, обитающие-на
глубинах, не
228
утилизируют
пищевые ресурсы приповерхностных вод,
и сущест-
вование
личинок резко расширяет кормовую базу
вида.
Для
ряда гидробионтов характерна
периодичность (прерывис-
тость)
развития. Как правило, она сопряжена
с циклическими из-
менениями
среды, когда благоприятные для развития
условия сме-
няются
неблагоприятными. Во время отливов
выключаются из ак-
тивной
жизни многие ракообразные, моллюски,
черви и другие оби-
татели
литорали. В водоемах средних и высоких
широт развитие
многих
гидробионтов прекращается в холодное
время года. Зака-
пываясь
в грунт высохших водоемов, приостанавливают
рост и
развитие
многие ракообразные, моллюски, некоторые
рыбы. Диа-
пауза
(перерыв в развитии) характерна для
ряда низших ракооб-
разных
и других гидробионтов при ухудшении
кислородных, тем-
пературных
и других условий.
Продолжительность.
Завершение развития организмов тесно
свя-
зано
с достижением ими некоторой массы,
величина которой далеко
не
постоянна для разных особей вида и, в
частности, очень сильно
зависит
от температуры. С ее повышением развитие
завершается
при
меньшей массе и длится меньшее время.
Например, длитель-
ность
развития личинок Ск.
йогваИв
при 15, 20, 25 и 30°С составляет
соответственно
34, 25, 16 и 16 дней, масса образовавшихся
куко-
лок—
7,3; 5,2; 4,6 и 3,7 мг. Значительное сокращение
длительности
развития
с одновременным уменьшением конечной
массы особей
хорошо
прослеживается у морских моллюсков с
продвижением из
высоких
широт в низкие. То же характерно для
морского планкто-
на.
Даже при обилии пищи планктонты живут
в высоких широтах
дольше,
чем родственные формы в более теплых
водах, и достигают
больших
размеров, хотя растут несколько
медленнее.
Влиянием
температуры в какой-то мере объясняется
сезонная
и
локальная изменчивость размеров
гидробионтов. Моллюск ЗШдиа
раШа,
обитающий на разных широтах побережья
США, имел сле-
дующие
длительности развития и конечные
размеры:
Широта,
5 35°11' 41°45' 46°58' 53°20' 60°27'
Длительность
жизни, годы 5 9 9 10 16
Размер,
см 12,7 13,5 14,5 14,6 16,4
В
опытах полихета Оркгуо1госка
1аЬгошса
из Неаполитанского
залива
при 16° С завершала цикл развития в 5
раз быстрее, чем при
14°
С. Развитие рачка АсагИа
1опза
из Чесапикского залива (США)
с
повышением температуры от 15,5 до 22,4 и
25,5° С сокращается
с
13 до 9 и 7 дней.
Во
многих случаях сокращение сроков
развития с повышением
температуры
довольно хорошо описывается уравнением
Белерадека:
Д=а(<—К)ь,
где И
— длительность развития, I
— температура во
время
развития, К
— минимальная температура, при которой
воз-
можен
процесс (так называемый «биологический
0»), а
и Ь
— кон-
станты.
Этим уравнением хорошо аппроксимируется
длительность
развития
личинок насекомых, многих морских
ракообразных, их
отдельных
стадий (СогсеИ, МсЬагеп, 1970). Часто для
расчетов
229
Рис.
39. Количество времени, затрачиваемого
на дыхательную ун-
дуляцию
(сплошные линии) личинками хирономид
при разной
концентрации
кислорода (пунктирные линии):
1
— СЫгопотиз рЫтозиз. 2
— С1урШепй1рез
раИепз, 3
— ЗупШатеза
пшоза
сроков
развития используется правило «суммы
тепла». Оно выра-
жается
уравнением (I—^о)^=соп51,
где I
— температура в период
развития
(/)), 1о
— температурный порог развития
(«биологиче-
ский
0»). Например, у личинок СЫгопотиз
йогзаНз,
выращиваемых
при
15, 20, 25 и 30° С, сумма тепла, выражаемая
числом градусо-
дней,
равнялась соответственно 315, 350, 306 и
384, у СН.
р1итозиз
при
тех же температурах — 600, 595, 615 и 640.
Характерно увели-
чение
суммы тепла, когда температура уклоняется
от оптимальной.
Помимо
указанных способов расчета сроков
развития, сущест-
вуют
и другие, в частности уравнения
Вант-Гоффа, Аррениуса.
Каждый
способ оправдывает себя в тех или иных
конкретных слу-
чаях,
но ни один из них не является универсальным.
Биологически-
ми
средствами организмы в той или иной
мере ограничивают тем-
пературные
эффекты.
В
какой-то степени аналогично действию
пониженных темпера-
тур
влияние повышенных давлений. С
продвижением на значитель-
ные
глубины и возрастанием давления
вязкость плазмы, как и при
низких
температурах, увеличивается, скорость
развития снижается,
конечные
размеры тела возрастают (см. гл. 1).
В
сильной степени темп развития
определяется трофическими и
респираторными
условиями. При дефиците корма циклы
развития
растягиваются
на неопределенно долгое время, такая
же картина
может
наблюдаться в условиях гипоксии.
Например, во время лет-
ней
и зимней стагнации водоемов, когда
вблизи дна кислорода
практически
нет, обитающие здесь животные впадают
в аноксибиоз
(см.
гл. 6). Живущие в трубках личинки хирономид
для поддержа-
ния
в них нужного уровня оксигенации воды
создают ее ток унду-
ляцией
тела. Чем меньше кислорода у дна, тем
чаще включается
ундуляция
и меньшая часть времени остается на
питание, в резуль-
230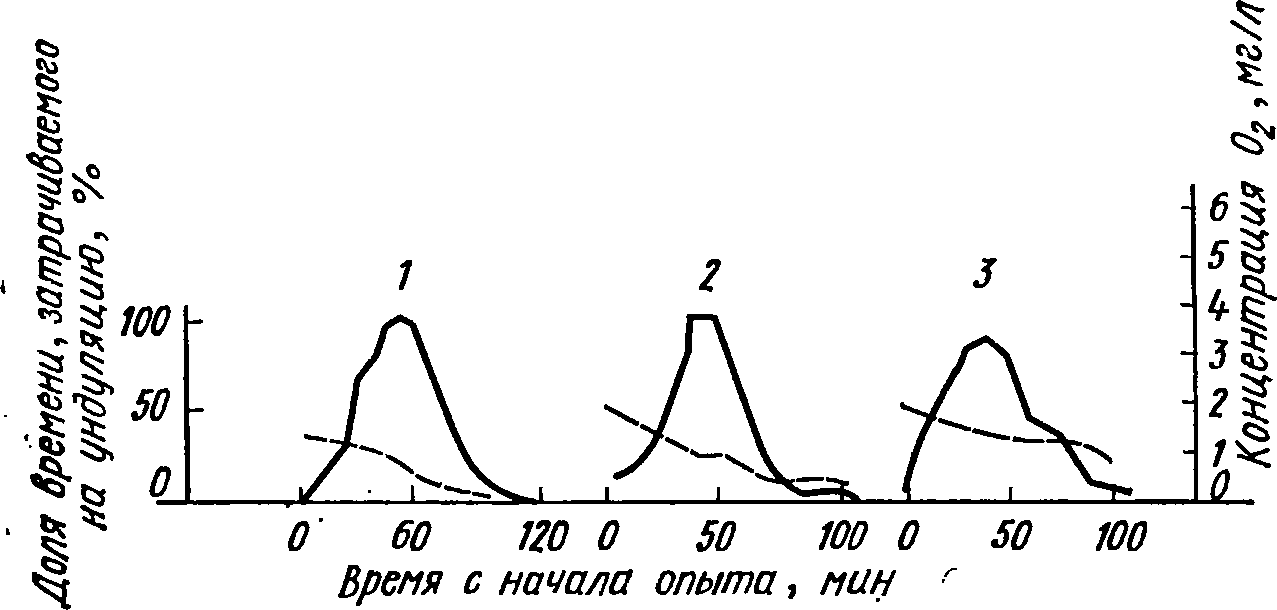
тате
чего замедляется развитие личинок
(рис. 39). Более или менее
сходная
картина наблюдается у многих других
обитателей трубок,
норок,
домиков и т. п.
Энергетика
роста и развития
Для
процессов биосинтеза, обеспечивающих
рост и развитие
особей,
необходимы энергетические траты, мерой
которых служит
интенсивность
газообмена. Соотношение величин
ассимиляции и
диссимиляции
характеризует эффективность трансформации
энер-
гии,
степень использования потребленной
пищи на рост. Величина
энтропии
энергии в процессе ее трансформации,
сильно варьиру-
ющая
в зависимости от видовых особенностей
организмов и условий
их
жизни, определяет КПД продукционных
процессов. Зная раци-
он,
усвояемость пищи, прирост, а также
величину катаболизма,
можно
достаточно точно судить об энергобалансе
особей, экономич-
ности
их роста. Эти знания — обязательное
условие рационального
освоения
природных экосистем и научной организации
аквакульту-
ры,
биологической оценки экономичности
выращивания тех или
иных
гидробионтов.
Интенсивность
трансформации энергии. Для особей
каждого ви-
да
та или иная удельная скорость
преобразования энергии отража-
ет
их генетические особенности и возможности
реализации биоло-
гических
потенций в конкретных условиях
окружающей среды. По-
этому
интенсивность трансформации энергии,
характеризующаяся
величиной
ее суммарного потока, отнесенной к
энергии, концентри-
рованной
в биомассе особи, сильно подвержена
влиянию меня-
ющихся
природных условий и неодинакова на
разных этапах онто-
генеза.
Показателем интенсивности преобразования
энергии может
служить
скорость оборота отдельных компонентов
тела, т. е. вели-
чина
их диссимиляции, отнесенная к содержанию
в биомассе. Пол-
ное
представление об интенсивности
трансформации энергии дает
сумма
ее энтропизации и аккумуляции за
единицу времени на еди-
ницу
биомассы (ее энергетического эквивалента).
В известной мере
скорость
преобразования энергии характеризуется
интенсивностью
ассимиляции
(см. гл. 4).
Как
правило, интенсивность трансформации
энергии с увеличе-
нием
размера (массы) особи снижается. Это
прослеживается как в
онтогенезе,
так и в ряду представителей разных
видов, различа-
ющихся
по массе тела. Бактерии, удваивающие
свою биомассу за
сутки,
за это время трансформируют количество
энергии, в 2—3 ра-
за
превышающее ее содержание в теле.
Несколько ниже темп пре-
образования
энергии бесцветными жгутиковыми и
другими пред-
ставителями
пикопланктона. Суточная трансформация
энергии
коловратками
и мелкими ракообразными обычно не
превышает
50—100%
от аккумулированной в теле, у более
крупных рачков она
составляет
15—20%, у червей и моллюсков—1—5%, у рыб,
как
правило,
выражается десятыми и сотыми долями
процента. По дан-
ным
Г. С. Карзинкина (1952), с увеличением
массы молоди бело-
231
рыбицы
от 32 до 260 и 860 мг интенсивность ассимиляции
снижалась
с
7 до 3 и 1,8% в сутки. У инфузорий туфелек,
питавшихся бакте-
риями
или хлореллой, интенсивность ассимиляции
соответственно
равнялась
16 и 75%, у дафний с повышением концентрации
пищи
(водорослей)
от 0,1 до 0,8; 1,5 и 4,8 интенсивность
ассимиляции
возрастала
с 0,8 до 5,9 и 13% в сутки (по углероду).
Суточная
потеря
углерода (0,4%), азота (0,2%) и фосфора (1,3%)
у криля,
обитающего
в холодных антарктических водах,
примерно на поря-
док
ниже, чем у представителей субтропического
и тропического
планктона
(1сес1а, МНсЬеИ, 1982).
Эффективность
использования пищи и энергии. Способ
оценки
эффективности
использования пищи на рост зависит от
целей ис-
следования.
В рыбоводстве и при выращивании водных
беспозво-
ночных
эффективность использования пищи на
рост часто выра-
жается
кормовым
коэффициентом,
под которым понимают отноше-
ние
массы корма, потребленного животными,
к их приросту вне за-
висимости
от химического состава пищи и тела
потребителя. Чем
выше
кормовой коэффициент, тем, следовательно,
хуже использу-
ется
корм на рост. Величина кормового
коэффициента у данного
животного
в очень большой степени зависит от
качества пищи.
Например,
для личинок хирономид, которых кормили
порошком из
макрофитов
и сухими дрожжами, он равнялся
соответственно 7
и
0,5. Если дрожжи даются не в сухом, а в
сыром виде, кормовой ко-
эффициент
возрастает в 3—4 раза. Следовательно,
он лишь в ма-
лой
степени характеризует свойства
организмов и мало отражает
степень
использования потребленных веществ
на рост. Например,
сухие
дрожжи используются на рост несколько
хуже сырых, но
кормовой
коэффициент в первом случае в 3—4 раза
ниже, чем во
втором.
Чтобы снять влияние воды, содержащейся
в пище, на по-
казатель
ее использования, вычисляют коэффициент
продуктивного
действия
корма
— отношение количества съеденной пищи
к приро-
сту
организма, причем обе величины выражают
в сухой массе
(Карзинкин,
1952). Иногда вычисляют отношение азота,
потреблен-
ного
с пищей, к его накоплению в организме.
В. С.
Ивлев (1938) предложил оценивать
эффективность ис-
пользования
пищи на рост двумя трофическими
коэффициентами.
Первый
из них (Ку)
определяется по уравнению /С1
= <21-’<2, где
(2!
— энергия вновь образованного в
организме вещества и (2
—
энергия
потребленной пищи. /Сг выражается
соотношением /Сг—
=
&1
■ №—@2),
где ф2
— энергия неусвоенной части пищи (С},
С} 1
и
<32
выражаются в единицах энергии, К\
и К2
— в процентах).
Величина
трофических коэффициентов видоспецифична,
зависит
от
состояния популяции и внешних условий.
Так, у планктонных
бактерий,
использующих РОВ, величина К\
летом в дельте Волги
достигала
60%, у аэробных и анаэробных целлюлозных
бактерий —
соответственно
3 и 1,2% (Горбунов, 1976). Для ряда
микрофла-
геллят
К\
и /С2
оказались равными 30 и 60% (РепсЬе1, 1982),
у аме-
бы
АтоеЬа
рго1еиз
при 10—11° С К\
составлял 4—29%, а при 15—■
20°
С лежал в пределах 16—47 %; в этих же
условиях К2
достигал со-
232
ответственно
49 и 65—82%
(Кодегзоп,
1881). У моллюс-
ков
5ркаегота
зиеысит
и
РьзйНит
сгаззит Кг
за вре-
мя
выращивания равнялся
44
и 29%.
С
возрастом коэффици-
ент
Кг
снижается, постепен-
но
падая до 0 по мере при-
ближения
размеров тела к
дефинитивным
(рис. 40).
Так,
у моллюсков 5. зиес1-
сит
с увеличением возраста
с
30 до 90 и 150 суток Кг
со-
ответственно
снижался с 65
до
35 и 15%, т. е. коэффици-
ент
использования пищи сни-
жался
в 2—3 раза, у моллюс-
ков
Р.
сгаззит
при таком же
увеличении
возраста Кг
сни-
жался
с 45 до 19 и 6% (Али-
мов,
1970). У голодных жи-
вотных
использование асси-
милированной
пищи на рост
выше,
чем у сытых. Например, с увеличением
длительности голода-
ния
мальков щуки от 0 до 10 и 20 суток величина
Кг
повышалась с
43
до 47 и 50%. Коэффициент Кг
возрастает с улучшением условий
питания,
когда на добывание пищи тратится меньше
энергии. Так,
с
повышением концентрации водорослей
с 0,2 до 0,5 и 1 млн. кле-
ток
на 1 мл Къ
у рачка Аг1ет1а
заИпа
возрастает с 24 до 26 и 28%.
Резко
снижается Кг
с ухудшением условий дыхания, с
повышением
температуры,
когда траты на дыхание возрастают
сильнее, чем при-
росты.
Коэффициенты
использования энергии хорошо отражают
осо-
бенности
ее трансформации безотносительно к
качеству потреблен-
ной
пищи и новообразованных веществ. В
некоторых аспектах, в
частности
в сфере практики, такой подход
недостаточен и пред-
ставление
об энергетике роста должно дополняться
знаниями осо-
бенностей
превращения веществ в популяциях.
Трансформация ве-
ществ
в процессе роста может идти таким
образом, что даже при
высоком
КПД окажется невыгодной с хозяйственной
точки зрения.
Если,
например, усваиваемая энергия будет
концентрироваться в
растущих
организмах в форме неусвояемых
высококалорийных ве-
ществ
(хитин и т. п.) или других малоценных
продуктов, то преоб-
разование
веществ хозяйственно неэффективно. С
другой стороны,
организмы,
использующие, пусть даже с низким КПД,
малодоступ-
ные,
трудноперевариваемые вещества
(клетчатка, гумус и т. п.) и
преобразующие
их в хозяйственно ценные продукты,
представляют
большой
практический интерес.
233-
I
*
\
3
1
I
«5
■О
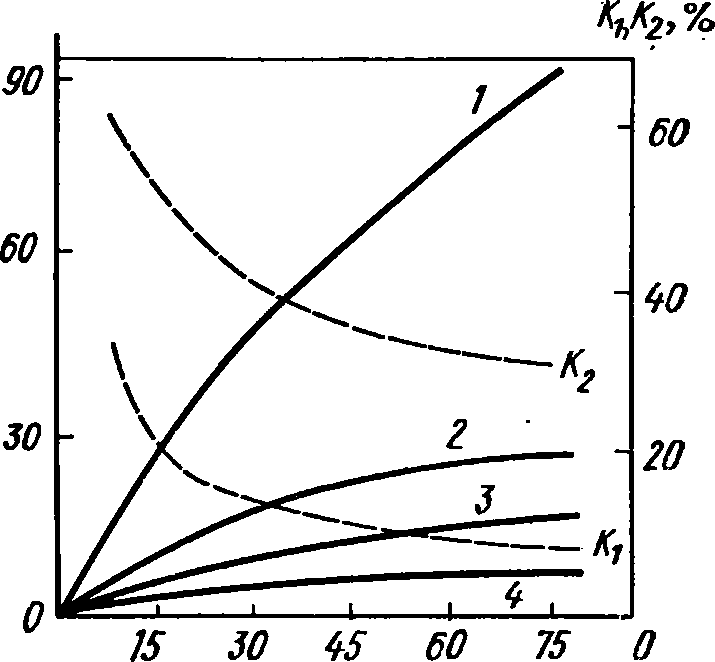
Масса, мг
Рис. 40. Элементы суточного энергобаланса рачка ОгсНевИа ЬоИае (по Сущене, 1967).
Энергия: 1 — потребленная, 2 — ассимилиро- ванная, 3 — рассеиваемая, 4 — аккумулированная в приросшей биомассе. /С» и /Сэ — коэффициенты использования энергии пищи иа рост
Одна
из характеристик использования пищи
— степень ее усвое-
ния
(переваривания), т. е. отношение
ассимилированной энергии и
потребленной.
У растительноядных форм этот показатель
заметно
ниже,
чем у плотоядных. Степень ассимиляции
потребленной пищи
сильно
меняется в зависимости от ее качества,
физиологических
особенностей
потребителей и условий питания.
Энтропизация
энергии. Жизнедеятельность гидробионтов,
как
и
всех организмов, сопровождается
рассеянием (деградацией) энер-
гии,
интегральной характеристикой которого
могут служить ско-
рость
и интенсивность потребления кислорода
в процессе дыхания.
На
окисление одинаковых количеств
белков, жиров и углеводов
тратится
разное количество кислорода; оно
почти точно соответст-
вует
количеству выделяющейся при этом
энергии: |
|
Теплотворная |
Затраты Ог на |
|
|
|
способность, |
сжигание 1 мг |
к |
|
|
Дж1мг (К) |
вещества (О), мг |
о |
Белки . . . |
|
24,2 |
1,75 |
13,8 |
Жиры . . . |
|
39,6 |
2,88 |
13,7 |
Углеводы . . |
, . . • • |
17,0 |
1,18 |
14,7 |
В
процессе биологического окисления на
единицу потребленного
кислорода
выделяется практически одно и то же
количество энер-
гии.
Следовательно, зная величину потребления
кислорода организ-
мами,
можно получить достаточно точное
представление об их
энергетических
тратах. Отношение количества энергии
(в джоулях),
выделяющейся
при окислении вещества, к массе
потребленного кис-
лорода
(в мг) получило название оксиэнергетического
(оксикало-
рийного)
коэффициента.
Его среднее значение для водных
организ-
мов
равно 14 Дж/мг Ог. Отдельные данные
уклоняются от найден-
ного
среднего значения только в третьем
знаке. Таким образом, из-
меряя
дыхание гидробионтов в разных условиях,
можно харак-
теризовать
их энергетику без непосредственного
учета отдельных
слагаемых
баланса. Величина оксиэнергетического
коэффициента
не
зависит от условий, в которых идет
аэробное дыхание.
Изучение
газообмена позволяет составить
представление не
только
о величине энергетических затрат у
гидробионтов, но и о том,
за
счет каких веществ добывается энергия,
т. е. судить о химиче-
ском
составе пищи. Такое суждение можно
получить, сравнивая
количество
потребляемого организмами Ог и
выделяемого ими СОг.
Когда
энергия добывается за счет питания
углеводами, организм
выделяет
столько СОг, сколько потребляет 02
(по объему). При
белковом
питании отношение выделяемого С02
к потребленному Ог
составляет
0,77—0,82, а в случае потребления жира —
0,7. Отноше-
ние
объема выделившегося в процессе дыхания
С02
к объему по-
требленного
02
получило название дыхательного
коэффициента.
Иногда
он превышает 1, что обычно обусловливается
частично ана-
эробным
добыванием энергии, когда организмы
испытывают недо-
статок
в кислороде. Так, при значительном
снижении концентра-
ции
02
дыхательный коэффициент червя ЗьрипсиШв
может возра-
стать
болёе чем в 14 раз:
234
Концентрация
Ог, мг/л 11,46 8,8 5,4 1,36 1,13 0,81
Дыхательный
коэффициент 0,69 1,03 Г,48 3,40 8,6 10,0
При
полном анаэробиозе дыхательный
коэффициент становится
величиной
бесконечной. На величину дыхательного
коэффициента
могут
также влиять температура и некоторые
другие факторы. Важ-
ной
характеристикой особенностей метаболизма
особей служит от-
ношение
потребленного кислорода к выделенному
азоту. Когда оно
мало
(<24), метаболизм идет преимущественно
за счет белка, если
велико
— источником энерготрат в основном
служат липиды. Для
гидробионтов
более характерен первый случай.
Например, для
криля
ЕирНазьа
зирегЬа
рассматриваемое отношение равно
28
(Зедаша
е1 а11., 1982) .Сходное значение оно имеет
у рыб.
Энергобаланс
особей. Длительное активное существование
гид-
робионтов
возможно, если получаемая ими энергия
больше рассеи-
ваемой,
т. е. энергобаланс положителен. Иногда
расход энергии
превышает
ее поступление и тогда наблюдается
отрицательный
рост
(снижение энергоемкости тела). Чем
стабильнее условия пи-
тания,
тем ниже устойчивость особей к голоданию.
По этой причи-
не
обитатели средних широт отличаются
наибольшей выносли-
востью
к продолжительной трате энергии,
превышающей ее поступ-
ление.
Более выровнен энергобаланс у
гидробионтов низких широт,
где
сезонные изменения кормовой базы
выражены слабее.
Положительная
часть энергобаланса особей складывается
из
энергии
веществ, поступающих перорально и
другими путями, в ча-
стности
осмотически. Эта энергия аккумулируется
в самой особи
(прирост),
расходуется на выполнение внешней
(передвижение, со-
здание
токов воды, рытье норок, построение
домиков и др.) и внут-
ренней
работы (пищеварение, выделение,
осморегуляция, нервные
импульсы
и др.). Часть ассимилированной энергии
отчуждается в
виде
половых продуктов, метаболитов и
структур, возобновляемых
в
процессе физиологической или репаративной
регенерации (пан-
цири,
экзувии, перитрофические мембраны,
чешуя и др.), а также
при
образовании разного рода домиков,
чехликов, паутинной вы-
стилки.
Существенна энергия, изымаемая
паразитами; некоторая
часть
потребленной пищи не усваивается,
выделяясь вместе с фека-
лиями.
Суммарное выражение энергобаланса
можно представить
уравнением
(Константинов, 1972)
Эр=Эв-|-Э8-|-Э1|-|-Эм-|-Э0-{-Эи-|-Эн, (5)
где
в левой части — энергия, поступившая
в организм (энергия ра-
циона),
а в правой — аккумулированная в теле
(Эа),
энтропизира-
ванная
(Ээ),
содержащаяся в выметываемых половых
продуктах
(Эп),
метаболитах (Эм),
отчуждаемых структурах (Э0),
изымаемая
паразитами
(Э„), неусвоенная (Эн).
Учет
и оценка всех слагаемых уравнения
сопряжена со многи-
ми
трудоемкими исследованиями. Поступаясь
некоторой точностью
расчетов,
используют более простую формулу
(Винберг, 1956):
235
р=п+т+н,
(6)
где
в левой части уравнения — энергия
рациона, в правой — энер-
гия
прироста (П), трат на дыхание (Т) и
неусвоения (Н). Вели-
чина
энерготрат оценивается по количеству
потребленного кислоро-
да,
т. е. учитывает всю внешнюю и внутреннюю
работу организма.
В
настоящее время в правую часть
балансового равенства (6)
обычно
включают и энергию выметываемых половых
продуктов.
Будучи
удобной расчетной схемой, балансовое
равенство (6) в
некоторых
случаях достаточно приемлемо описывает
энергобаланс
особей.
Чаще статьи уравнения (5), не учитываемые
балансовым
равенством
(6), настолько существенны, что их
игнорирование в
целях
простоты расчетов приводит к большой
неточности. Напри-
мер,
совсем не ничтожна энергия выделяемых
метаболитов, в част-
ности
аммиака и других недоокисленных
продуктов белкового об-
мена,
на долю которых может приходиться
более 10—15% энергии
рациона
(Бретт, Гроувс, 1983). Неточность балансового
равенства
(6)
усугубляется тем, что под рационом
понимают то количество
пищи,
которое животные воспринимают
перорально. Однако нема-
ловажное
значение в энергетике гидробионтов
имеет осмотическое
питание.
Например, часовой рацион бесцветного
жгутиконосца
РагаЬойо
аНепиаЫз
в среднем составлял 1Ы0~8Дж
или 50%
энергии
тела, причем потребление РОВ (осмотическое
питание)
полностью
компенсировало все затраты на дыхание.
Сопоставление
уравнений (5) и (6) позволяет уточнить
смысл
трофических
коэффициентов К\
и Кг-
Очевидно, они отражают сте-
пень
утилизации пищи на рост, но не
эффективность трансформа-
ции
энергии организмом. Для оценки КПД
трансформации энергии
можно
использовать коэффициенты /С/ и Кг
(Заика, 1974):
К[=(А—
Т)/Р;
Кг=(А-Т)/(Р-Н).
Поскольку
разница между энергией ассимилированной
(А)
и
рассеиваемой
(Т)
выше, чем энергия прироста ((21),
коэффициенты
трансформации
энергии (К\
и Кг')
всегда выше трофических
(Ку
и Кг)
-
Как показал В. Е. Заика, они возрастают
с уменьшением
сроков
жизни гидробионтов и у короткоживущих
организмов при-
ближаются
к 70—80%.
Соотношение
величин отдельных слагаемых в
энергобалансе у
разных
гидробионтов сильно варьирует в
зависимости от их видо-
вой
принадлежности, физиологического
состояния и внешних ус-
ловий.
Например, у рачка ЕирНаизьа
расЦьса
при 8 и 12° С исполь-
зуется
на рост соответственно 9,6 и 9,7%
ассимилированной энер-
гии
(углерода), расходуется на линьку 3 и
3,6%, на размножение —
1
и 11,5% (Кобз, 1982). Интересно, что в данном
случае различие
Температур
почти не сопровождается сдвигами в
энергобалансе, хо-
тя
длительность развития сокращается с
336 суток в первом случае
до
210 во втором. Максимальные значения
К\
у ' беспозвоночных
обычно
лежат в диапазоне 30—40%, снижаясь с
возрастом. Вели-
чины
Кг
часто достигают 60—70%, но в большинстве
случаев не
превышают
40—50%.