

Потенциал концевой пластинки
В нервно-мышечном синапсе ацетилхолин синтезируется в окончанияхдвигательных нервов и накапливается в пузырьках. Когда в окончание приходит потенциал действия, ацетилхолин из 150-200 пузырьков высвобождается в синаптическую щель и связывается с холинорецепторами (холинорецепторы нервно-мышечных синапсов принадлежат кN-холинорецепторам ), плотность которых особенно высока на гребнях складок постсинаптической мембраны. Каналы, сопряженные с холинорецепторами, открываются, в клетку входят катионы (в основном Na+), и происходит деполяризация постсинаптической мембраны, называемая потенциалом концевой пластинки. Поскольку этот потенциал в норме всегда сверхпороговый, он вызывает потенциал действия, распространяющийся по мышечному волокну и вызывающий сокращение. Потенциал концевой пластинки короткий, так как ацетилхолин, во-первых, быстро отсоединяется от рецепторов, во-вторых - гидролизуется АХЭ .
-
Классификация нервных волокон. Распространения возбуждения по безмиелиновым и миелиновым нервным волокнам. Характеристика их возбудимости и лабильности. Законы проведения возбуждения по нерву.
Наиболее распространена классификация по Дж.Эрлангеру и Х.Гассеру (1937), в которой волокна разделяют на три типа: А, В и С (табл.3). Волокна типа А и В являются миелиновыми, типа С — безмиелиновыми. Волокна А делят на 4 подгруппы: α, β, γ, δ. В периферической нервной системе к волокнам Аα относятся афферентные волокна от механорецепторов кожи, мышечных и сухожильных рецепторов, а также эфферентные волокна к скелетным мышцам. К Аβ принадлежат афферентные волокна от кожных рецепторов прикосновения и давления, от части мышечных и висцеральных рецепторов. Аγ представляют собой эфферентные волокна, через которые регулируется активность мышечных рецепторов. К Аδ относят афферентные волокна от части тактильных, температурных и болевых, а также суставных рецепторов. К волокнам типа В принадлежат преганглионарные волокна вегетативной нервной системы. К волокнам типа С относят постганглионарные волокна вегетативной нервной системы, афферентные волокна от некоторых болевых (вторичная боль), тепловых и висцеральных рецепторов.
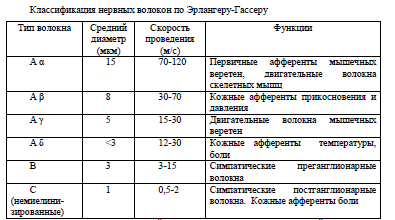
Из данных, представленных в таблице, видно, что средний диаметр каждого типа волокна снижается от типа А до С (каждый примерно в 2 раза по отношению к предыдущему). Соответственно этому снижается и скорость проведения возбуждения. Низкая скорость проведения нервного импульса в волокнах типа С связана с особенностями проведения возбуждения в безмиелиновых волокнах. Лабильность также уменьшается от волокон Аα до С и находится в обратной зависимости от продолжительности фазы абсолютной рефрактерности. Возбудимость тоже уменьшается от волокон Аα (наибольшая возбудимость) к волокнам С (наименьшая возбудимость). Например, пороговая сила электрического тока у волокон С в 30—50 раз больше, чем у волокон Аα. Исследование факторов, блокирующих нервную проводимость, показало, что к давлению наиболее чувствительны волокна А, к кислородному голоданию (гипоксии) — волокна В, к местным анестетикам — волокна С.
Проведение импульса по нервным волокнам.
ПД или нервный импульс может возникать в любой точке возбудимой мембраны нервного или мышечного волокна и способен распространяться вдоль ее поверхности. При этом роль ПД заключается в передаче информации по нервным волокнам от тела нейрона к нервному окончанию. Когда ПД достигают терминалей аксона, то информация благодаря выделению из нервных окончаний молекул медиатора передается на другие нейроны. В мышечных клетках ПД распространяются по сарколемме и активируют механизм сокращения мышц. Проведение НИ от тела нейрона к окончанию аксона различается в немиелинизированных и миелинизированных нервных волокнах.
Немиелинизированные волокна.
Проведение ПД по ННВ происходит путем активации потенциалзависимых натриевых ионных каналов участка мембраны волокна, прилегающего к тому месту, где возник ПД. При этом между возбужденным и невозбужденным участками мембраны нервного волокна возникают локальные электрические токи, вызывающие деполяризацию мембраны невозбужденного участка до критического уровня. После этого в мембране НВ мгновенно открываются потенциалзависимые натриевые каналы. ПД, т.о., генерируется в последующем участке НВ.
Миелинизированные волокна.
В МНВ ПД генерируются только в области перехвата Ранвье, т.е. той части мембраны, которая не покрыта шванновскими клетками. С одной стороны, это обусловлено тем, что цитоплазма шванновской клетки содержит липид – сфингомиелин, который уменьшает поток ионов через мембрану НВ примерно в 5000 раз и снижает ее емкость в 50 раз. С другой стороны, в области ПР в мембране НВ имеется наибольшее число потенциалзависимых натриевых ионных каналов, а сама мембрана обладает нормальным уровнем возбудимости. При нанесении точечного удара на МНВ ПД генерируется в зоне ПР, и возникают электрические токи, которые текут вдоль силовых линий от плюса к минусу потенциала на мембране. Одновременно в аксоплазме НВ возникают продольные токи, которые направлены от места генерации ПД в обе стороны. Продольные токи вызывают открывание ПЗНИК, а следовательно, движение ионов Na+ через мембрану и генерацию ПД в соседних ПР, минуя часть НВ, покрытого шванновскими клетками. Поскольку ПД передаются (перепрыгивают) от одного ПР к другому, то механизм проведения ПД в МНВ получил название сальтаторного. В МНВ реполяризация мембраны после ПД происходит с очень высокой скоростью. В результате НВ имеют высокую функциональную лабильность и способны проводить значительное число ПД в единицу времени. Скорость распространения ПД в зависимости от диаметра и типа МНВ чрезвычайно высока и варьирует от 6 до 120 м /с. В толстых МНВ скорость проведения возбуждения пропорциональна диаметру волокна, а проводимость НВ имеет обратную зависимость от его диаметра.
Законы проведения возбуждения по нервному волокну:
-
Закон физиологической непрерывности – проведение возбуждения по НВ возможно при условии его структурной целостности и физиологической непрерывности. Физиологическая непрерывность нерва м.б. нарушена, например, при сдавлении нерва без его структурного повреждения, что препятствует проведению ПД.
-
Закон изолированного проведения – при проведении возбуждения по НВ ПД не распространяется с одного волокна на другое, например, рядом расположенное.
-
Закон двухстороннего проведения – отдельно нервное волокно обладает двухсторонней проводимостью. Так, при искусственном электрическом раздражении в любой точке по ходу НВ может возникать ПД и распространяться как центростремительно, так и центробежно.
Лабильность. Парабиоз и его фазы (Н.Е.Введенский).
-
Лабильность — функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной тканях. Понятие "Л." введено русским физиологом Н. Е. Введенским (1886), который считал мерой Л. наибольшую частоту раздражения ткани, воспроизводимую ею без преобразования ритма. Л. отражает время, в течение которого ткань восстанавливает работоспособность после очередного цикла возбуждения. Наибольшей Л. отличаются отростки нервных клеток — аксоны, способные воспроизводить до 500—1000 импульсов в 1 сек; менее лабильны центральные и периферические места контакта — синапсы (например, двигательное нервное окончание может передать на скелетную мышцу не более 100—150 возбуждений в 1 сек). Угнетение жизнедеятельности тканей и клеток (например, холодом, наркотиками) уменьшает Л., т. к. при этом замедляются процессы восстановления и удлиняется рефрактерный период.
Парабиоз — состояние, пограничное между жизнью и смертью клетки.
Причины парабиоза – самые разные повреждающие воздействия на возбудимую ткань или клетку, не приводящие к грубым структурным изменениям, но в той или иной мере нарушающее ее функциональное состояние. Такими причинами могут быть механические, термические, химические и другие раздражители.
Сущность парабиоза. Как считал сам Введенский, в основе парабиоза лежит снижение возбудимости и проводимости, связанное с натриевой инактивацией. Советский цитофизиолог Н.А. Петрошин полагал, что в основе парабиоза лежат обратимые изменения белков протоплазмы. Под действием повреждающего агента клетка (ткань), не теряя структурной целостности, полностью прекращает функционировать. Это состояние развивается фазно, по мере действия повреждающего фактора (то есть зависит от продолжительности и силы действующего раздражителя). Если повреждающий агент вовремя не убрать, то наступает биологическая смерть клетки (ткани). Если же этот агент убрать вовремя, то ткань так же фазно возвращается в нормальное состояние.
Эксперименты Н.Е. Введенского.
Введенский проводил опыты на нервно-мышечном препарате лягушки. На седалищный нерв нервно-мышечного препарата последовательно наносились тестирующие раздражители разной силы. Один раздражитель был слабый (пороговой силы), то есть вызывал минимальное по величине сокращение икроножной мышцы. Другой раздражитель был сильный (максимальный), то есть наименьший из тех, которые вызывают максимальное сокращение икроножной мышцы. Затем в какой-либо точке на нерв наносился повреждающий агент и каждые несколько минут нервно-мышечного препарат подвергался тестированию: поочередно слабыми и сильными раздражителями. При этом последовательно развивались следующие стадии:
-
Уравнительная, когда в ответ на слабый раздражитель величина сокращения мышцы не изменялась, а в ответ на сильный амплитуда сокращения мышцы резко уменьшалась и становилась такой же, как при ответе на слабый раздражитель;
-
Парадоксальная, когда в ответ на слабый раздражитель величина сокращения мышцы оставалась прежней, а в ответ на сильный раздражитель величина амплитуды сокращения становилась меньше, чем в ответ на слабый раздражитель, или мышца вообще не сокращалась;
-
Тормозная, когда и на сильный и на слабый раздражители мышца не отвечала сокращением. Именно это состояние ткани и обозначается как парабиоз.
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
1. Нейрон как структурная и функциональная единица ЦНС. Его физиологические свойства. Строение и классификация нейронов.
Нейроны– это основная структурно-функциональная единица нервной системы, обладающая специфическими проявлениями возбудимости. Нейрон способен принимать сигналы, перерабатывать их в нервные импульсы и проводить к нервным окончаниям, контактирующим с другим нейроном или рефлекторными органами (мышца или железа).
Виды нейронов:
-
Униполярные (имеют один отросток – аксон; характерны для ганглиев беспозвоночных);
-
Псевдоуниполярные (один отросток, делящийся на две ветви; характерно для ганглиев высших позвоночных).
-
Биполярные (есть аксон и дендрит, характерно для периферических и чувствительных нервов);
-
Мультиполярные (аксон и несколько дендритов – характерно для мозга позвоночных);
-
Изополярные (трудно дифференцировать отростки би- и мультиполярных нейронов);
-
Гетерополярные (легко дифференцировать отростки би- и мультиполярных нейронов)
Функциональная классификация:
1.Афферентные (чувствительные, сенсорные – воспринимают сигналы из внешней или внутренней среды);
2.Вставочные связывающие нейроны друг с другом (обеспечивают передачу информации внутри ЦНС: с афферентных нейронов на эфферентные).
3. Эфферентные (двигательные, мотонейроны – передают первые импульсы от нейрона к исполнительным органам).
Главная структурная особенностьнейрона – наличие отростков (дендритов и аксонов).
1–
дендриты;
2 – тело клетки;
3 – аксонный холмик;
4 – аксон;
5 –Швановская клетка;
6 – перехват Ранвье;
7 – эфферентные нервные окончания.
Последовательное синоптическое объединение всех 3х нейронов образует рефлекторную дугу.
Возбуждение, возникшее в виде нервного импульса на каком-либо участке мембраны нейрона, пробегает по всей его мембране и по всем его отросткам: как по аксону, так и по дендритам.Передаётсявозбуждение от одной нервной клетки к другойтолько в одном направлении- с аксонапередающегонейрона навоспринимающийнейрон черезсинапсы, находящиеся на его дендритах, теле или аксоне.
Одностороннюю передачу возбуждения обеспечивают синапсы. Нервное волокно (отросток нейрона) может передавать нервные импульсыв обоих направлениях, а односторонняя передача возбуждения появляется тольков нервных цепях, состоящих из нескольких нейронов, соединённых синапсами.Именно синапсы обеспечивают одностороннюю передачу возбуждения.
Нервные клетки воспринимают и перерабатывают поступающую к ним информацию. Эта информация приходит к ним в виде управляющих химических веществ:нейротрансмиттеров. Она может быть в видевозбуждающихилитормозныххимических сигналов, а также в видемодулирующихсигналов, т.е. таких, которые изменяют состояние или работу нейрона, но не передают на него возбуждение.
Нервная система играет исключительную интегрирующую рольв жизнедеятельности организма, так как объединяет (интегрирует) его в единое целое и интегрирует его в окружающую среду. Она обеспечивает согласованную работу отдельных частей организма (координацию), поддержание равновесного состояния в организме (гомеостаз) и приспособление организма к изменениям внешней или внутренней среды (адаптивное состояниеи/илиадаптивное поведение).
Нейрон — это нервная клетка с отростками, являющаяся основной структурной и функциональной единицей нервной системы. Она имеет строение, сходное с другими клетками: оболочка, протоплазма, ядро, митохондрии, рибосомы и другие органоиды.
В нейроне различают три части: тело клетки — сома, длинный отросток — аксон и множество коротких разветвленных отростков — дендритов. Сома выполняет обменные функции, дендриты специализируются на приеме сигналов из внешней среды или от других нервных клеток, аксон на проведении и передаче возбуждения в область, удаленную от зоны дендритов. Аксон оканчивается группой концевых разветвлений для передачи сигналов другим нейронам или органам-исполнителям. Наряду с общим сходством в строении нейронов наблюдается большое разнообразие, обусловленное их функциональными различиями (рис. 1).
ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА НЕЙРОНОВ
Основные свойства нейронов: раздражимость, возбудимость, проводимость, лабильность, инертность, утомляемость, торможение, регенерация и др.
Раздражимость — способность нервной клетки отвечать на различные раздражения биохимическими изменениями, сопровождающимися нарушением ионного равновесия и деполяризацией электрических зарядов на мембранах клетки в месте раздражения. Раздражимость присуща всем клеткам, и особенно нервным, связанным с чувствительным восприятием запаховых, звуковых, световых и других раздражителей. Раздражимость — пусковой механизм проявления другого свойства — возбудимости.
Возбудимость — способность отдельных частей нервной клетки генерировать электрохимические импульсы, т. е. отвечать на раздражение возбуждением. Для перехода нервной клетки в состояние возбуждения необходимо, чтобы сила действующего раздражителя достигла критического предела — пороговой величины. Способность нейрона отвечать возбуждением на наименьшую силу раздражителя называется нижним порогом возбудимости. Чем чувствительнее нервная клетка к раздражению, тем меньше порог возбудимости, и, следовательно, даже самый слабый раздражитель может вызвать возбуждение. Величина возбуждения нейрона зависит от силы раздражителя и возрастает по закону силовых отношений до определенного предела — верхнего порога возбудимости. Применение раздражителей сверхпороговой силы создает в нейроне запредельное торможение, которое охраняет нервную клетку от перевозбуждения (рис. 2).
Одиночное раздражение обычно вызывает серию импульсов определенной силы, продолжительности и частоты. В разных нервных клетках частота импульсов различная — от 100 до 1000 в секунду. Сила и продолжительность импульсов возбуждения зависит от характера раздражения.
Проводимость — способность нейрона проводить импульсы возбуждения с определенной скоростью, в неизменном ритме и силе. Возбуждение по нервному волокну может распространяться в обе стороны от раздражаемого участка. В разных нервных клетках скорость проведения возбуждения неодинакова и зависит от физиологического состояния нейрона и толщины волокна. В чувствительных нейронах возбуждение распространяется со скоростью 100–120 метров в секунду, в двигательных — 60–100, а в вегетативной нервной системе — 5–7.
Лабильность (подвижность) — способность нервной клетки принимать и передавать максимальное число импульсов за единицу времени без искажения. Подвижность двигательных нейронов не более 500 импульсов в секунду. Лабильность обеспечивает направленное распределение и проведение импульсов возбуждения нужной частоты по определенным нервным путям. В процессе роста и развития организма, а также при систематической тренировке, лабильность увеличивается и обеспечивает динамичность нервной системы, при утомлении и старении — уменьшается.
Инертность — способность нервной клетки накапливать и хранить в себе следы возбуждения и торможения. Полученная информация откладывается в дендритах, соме клетки, хромосомах ядра в виде биохимических изменений ДНК и РНК плазмы. Это свойство нейронов обеспечивает память организма, которая имеет решающее значение в процессе обучения животных.
Утомляемость — естественный процесс снижения работоспособности клетки при длительном возбуждении или торможении. Проявляется в виде уменьшения силы возбуждения, замедления частоты ритма импульсов и скорости их проведения. Отдых нервных клеток или смена нервной деятельности снимает утомление, и все свойства восстанавливаются.
Торможение — процесс, обратный возбуждению. Заключается в ослаблении, остановке или предупреждении возникновения возбуждения. Торможение — активный процесс, распространяясь по нервным клеткам, он обеспечивает согласованную работу отдельных органов и всего организма в целом.
Регенерация — способность нервной клетки восстанавливать утраченные или поврежденные отростки путем прорастания. Нервные клетки не размножаются, погибшие нейроны не восстанавливаются. Волокна нервной клетки способны прорастать, если сохранилось тело клетки.
2. Учение о рефлексе (Р.Декарт, Г.Прохазка), его развитие в трудах И.М.Сеченова, И.П.Павлова, П.К.Анохина. Классификация рефлексов. Рефлекторный путь, обратная афферентация и ее значение. Время рефлекса. Рецептивное поле рефлекса.
Рефлекс (от лат. reflexus — отражённый) — стереотипная реакция живого организма на раздражитель, проходящая с участием нервной системы. Принцип рефлекса – это универсальная и своеобразная форма взаимодействия организма со средой, происходящая при участии нервной системы. Понятие о рефлексе возникло в XVI веке в учении Р. Декарта (1596-1650) о механической картине мира. Под рефлексом Р. Декарт понимал движение «животных духов» от мозга к мышцам по типу отражения светового луча. Согласно его схеме внешние предметы действуют на периферические окончания расположенных внутри нервных «трубок» нервных «нитей», которые, натягиваясь, открывают клапаны отверстий, ведущих из мозга в нервы. По каналам этих нервов «животные духи» перемещаются в соответствующие мышцы, которые в результате раздуваются, и, таким образом, происходит движение.
Биологическая концепция рефлекса была сформирована чешским анатомом и физиологом Йиржи Прохазкой (1749-1820). Свои представления о рефлексе Й. Прохазка выразил следующим образом: внешние впечатления, возникающие в чувствительных нервах, быстро распространяются по всей их длине до самого начала. Там они отражаются по определенному закону, переходят на соответствующие им двигательные нервы и по ним очень быстро направляются к мышцам, которые затем производят точные и строго ограниченные движения. Впервые термин «рефлекс» был введен в научный язык Й. Прохазкой.
В дальнейшем, уже в XIX в., была создана рефлекторная теория нервной деятельности. Дуализм Р. Декарта в понимании рефлекторной природы деятельности нервной системы был преодолен И. М. Сеченовым, который в «Рефлексах головного мозга» (1863) впервые четко обосновал, что явления сознания подчиняются физиологическим законам и что в основе психических явлений лежат рефлекторные процессы.
В дальнейшем И. П. Павлов на примерах образования условных рефлексов показал, что поведение животных обусловлено рефлекторными механизмами. Механизмы поведения по И. П. Павлову основываются на трех принципах рефлекторной деятельности: принцип детерминизма (причинности) — всякое действие организма причинно обусловлено; принцип анализа и синтеза — любое воздействие вначале анализируется качественно, количественно, по биологической значимости, а затем в зависимости от результата анализа синтезируется соответствующее ответное поведение; принцип структурности — все физиологические процессы протекают в определенных нервных структурах.
По ряду признаков рефлексы могут быть разделены на группы:
-
По типу образования: условные и безусловные рефлексы
-
По видам рецепторов: экстероцептивные (кожные, зрительные, слуховые, обонятельные), интероцептивные (с рецепторов внутренних органов) и проприоцептивные (с рецепторов мышц, сухожилий, суставов)
-
По эффекторам: соматические, или двигательные (рефлексы скелетных мышц), например флексорные, экстензорные, локомоторные, статокинетические и др.; вегетативные внутренних органов — пищеварительные, сердечно-сосудистые, выделительные, секреторные и др.
-
По биологической значимости: оборонительные, или защитные, пищеварительные, половые, ориентировочные.
-
По степени сложности нейронной организации рефлекторных дуг различают моносинаптические, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный), и полисинаптические, дуги которых содержат также 1 или несколько промежуточных нейронов и имеют 2 или несколько синаптических переключений (например, флексорный).
-
По характеру влияний на деятельность эффектора: возбудительные — вызывающими и усиливающими (облегчающими) его деятельность, тормозные — ослабляющими и подавляющими её (например, рефлекторное учащение сердечного ритма симпатическим нервом и урежение его или остановка сердца — блуждающим).
-
По анатомическому расположению центральной части рефлекторных дуг различают спинальные рефлексы и рефлексы головного мозга.
Рефлекс - ответная реакция организма на раздражение из внешней или внутренней среды, осуществляющаяся при участии ЦНС. Вся нервная деятельность складывается из рефлексов различной степени сложности, т. е. является отраженной, вызванной внешним поводом, внешним толчком. Путь, по которому проходит нервный импульс от рецептора до эффектора (действующий орган), называется рефлекторной дугой.
В рефлекторной дуге различают пять звеньев: 1) рецептор; 2) чувствительное волокно, про-водящее возбуждение к центрам; 3) нервный центр, где происходит переключение возбуждения с чувствительных клеток на двигательные; 4) двигательное волокно, передающее нервные импульсы на периферию; 5) действующий орган - мышца или железа. Для осуществления любого рефлекса необходима целостность всех звеньев рефлекторной дуги. Нарушение хотя бы одного из них ведет к исчезновению рефлекса.
Любое раздражение, воспринимаемое рецептором, кодируется в нервный импульс и в таком виде по чувствительным волокнам направляется в ЦНС. Здесь эта информация перерабатывается, отбирается и передается на двигательные нервные клетки, которые посылают нервные импульсы к рабочим органам - мышцам, железам и вызывают тот или иной приспособительный акт - движение или секрецию.
Во время ответной реакции возбуждаются рецепторы рабочего органа и от них в ЦНС поступают импульсы - информация о достигнутом результате. Живой организм, как любая саморегулирующаяся система, работает по принципу обратной связи. Афферентные импульсы, осуществляющие обратную связь, либо усиливают и уточняют реакцию, если она не достигла цели, либо прекращают ее. Таким образом, рефлекс осуществляется не рефлекторной дугой, а рефлекторным кольцом; рефлекс заканчивается по достижении результата.
Время рефлекса.
Время, прошедшее от момента нанесения раздражения до ответа на него, называется временем рефлекса (латентный период). Оно слагается из времени, необходимого для возбуждения рецепторов, проведения возбуждения по чувствительным волокнам, ЦНС, двигательным волокнам, и, наконец, скрытого (латентного) периода возбуждения рабочего органа. Большая часть времени уходит на проведение возбуждения через нервные центры - центральное время рефлекса. Это объясняется тем, что в синапсах ЦНС происходит замедление проведения возбуждения, так называемая синаптическая задержка. Чем меньше нейронов входит в состав рефлекторной дуги, тем короче время рефлекса. Поэтому сухожильные рефлексы, возникающие при растяжении сухожилия, имеющие двухнейронную дугу, наиболее быстрые. Их время составляет всего 19-23 мс, тогда как время рефлекса моргания, возникающего при раздражении глаза, равно 50-200 мс. Наибольшим является время вегетативных рефлексов. Время рефлекса зависит от силы раздражения и возбудимости ЦНС.
Рецептивное поле рефлекса.
Каждый рефлекс можно вызвать только с определенного рецептивного поля. Анатомическая область, при раздражении которой вызывается данный рефлекс, носит название рецептивного поля рефлекса. Например, рефлекс сосания возникает при раздражении губ ребенка, рефлекс сужения зрачка - при освещении сетчатки, коленный рефлекс (разгибание голени) - при легком ударе по сухожилию ниже надколенника.