

4. Потенциал действия. Ионный механизм возникновения потенциала действия
Потенциалом действия (ПД) называют быстрое колебание мембранного потенциала, возникающее при возбуждении нервных, мышечных и некоторых других клеток. В его основе лежат изменения ионной проницаемости мембраны. Амплитуда ПД мало зависит от силы вызывающего его раздражителя, важно лишь, чтобы эта сила была не меньше некоторой критической величины, которая называется порогом раздражения. Возникнув в месте раздражения, ПД распространяется вдоль нервного или мышечного волокна, не изменяя своей амплитуды.
В естественных условиях ПД генерируются в нервных волокнах при раздражении рецепторов или возбуждении нервных клеток. Распространение ПД по нервным волокнам обеспечивает передачу информации в нервной системе. Достигнув нервных окончаний, ПД вызывают секрецию химических веществ (медиаторов), обеспечивающих передачу сигнала на мышечные или нервные клетки. В мышечных клетках ПД инициируют цепь процессов, вызывающих сократительный акт. Ионы, проникающие в цитоплазму во время генерации ПД, оказывают регулирующее влияние на метаболизм клетки и, в частности, на процессы синтеза белков, составляющих ионные каналы и ионные насосы.
В
ПД выделяют следующие фазы (рис. 3):
деполяризации;
реполяризации.
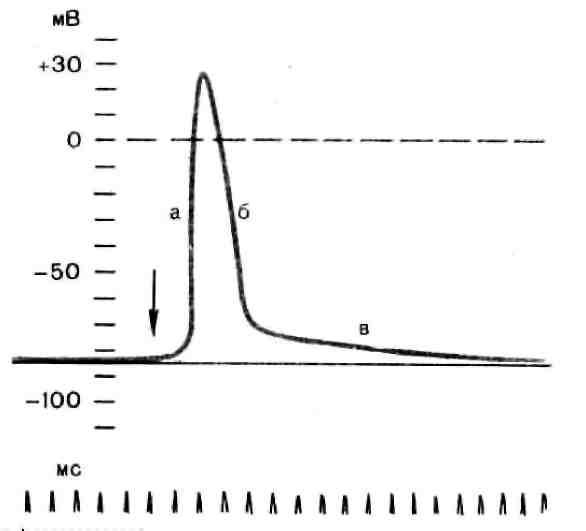
Рис. 3. Потенциал действия скелетного мышечного волокна, зарегистрированный с помощью внутриклеточного микроэлектрода: а – фаза деполяризации, б – фаза реполяризации, в – фаза следовой деполяризации (отрицательный следовой потенциал). Момент нанесения раздражения показан стрелкой.
Установлено, что во время восходящей фазы (фазы деполяризации) происходит не просто исчезновение потенциала покоя (как это первоначально предполагали), а возникает разность потенциалов обратного знака: внутреннее содержимое клетки становится заряженным положительно по отношению к наружной среде, иными словами, происходит реверсия мембранного потенциала. Во время нисходящей фазы (фазы реполяризации) мембранный потенциал возвращается к своему исходному значению. Если рассмотреть пример записи ПД в скелетном мышечном волокне лягушки (см. рис. 3), то видно, что в момент достижения пика мембранный потенциал составляет +30 – +40 мВ. Длительность пика ПД у различных нервных и мышечных волокон варьирует от 0,5 до 3 мс, причем фаза реполяризации продолжительнее фазы деполяризации.
Изменения мембранного потенциала, следующие за пиком потенциала действия, называют следовыми потенциалами. Различают два вида следовых потенциалов – следовую деполяризацию и следовую гиперполяризацию.
Ионный механизм возникновения ПД. Как отмечалось, в состоянии покоя проницаемость мембраны для калия превышает ее проницаемость для натрия. Вследствие этого поток К+ из цитоплазмы во внешний раствор превышает противоположно направленный поток Na+. Поэтому наружная сторона мембраны в покое имеет положительный потенциал по отношению к внутренней.
При действии на клетку раздражителя проницаемость мембраны для Na+ резко повышается и становится примерно в 20 раз больше проницаемости для K+. Поэтому поток Na+ из внешнего раствора в цитоплазму начинает превышать направленный наружу калиевый ток. Это приводит к изменению знака (реверсии) мембранного потенциала: внутренняя сторона мембраны в месте возбуждения становится заряженной положительно по отношению к ее наружной поверхности. Указанное изменение мембранного потенциала соответствует восходящей фазе ПД (фазе деполяризации).
Повышение проницаемости мембраны для Na+ продолжается лишь очень короткое время. Вслед за этим проницаемость мембраны для Na+ вновь понижается, а для K+ возрастает. Процесс, ведущий к понижению ранее увеличенной натриевой проницаемости мембраны, назван натриевой инактивацией. В результате инактивации поток Na+ внутрь цитоплазмы резко ослабляется. Увеличение же калиевой проницаемости вызывает усиление потока K+ из цитоплазмы во внешний раствор. В итоге этих двух процессов и происходит реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к наружной стороне мембраны. Этому изменению потенциала соответствует нисходящая фаза ПД (фаза реполяризации). Опыты на гигантских нервных волокнах кальмара позволили получить подтверждение правильности натриевой теории возникновения ПД.
ПД возникает при деполяризации поверхностной мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению ионов Na внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные изменения на мембране (локальный ответ). При увеличении силы раздражения, когда достигнут порог возбудимости, изменения мембранного потенциала достигают критического уровня деполяризации (КУД). Например, величина потенциала покоя равна -70 мВ, КУД = -50 мВ. Чтобы вызвать возбуждение, надо деполяризовать мембрану до -50 мВ, т.е. на -20 мВ снизить ее исходный потенциал покоя. Только при достижении КУД наблюдается резкое изменение мембранного потенциала, которое регистрируется в виде ПД. Таким образом, основное условие возникновения потенциала действия – это снижение мембранного потенциала до критического уровня деполяризации.
В основе рассмотренных изменений ионной проницаемости мембраны при генерации ПД лежат процессы открывания и закрывания специализированных ионных каналов в мембране, обладающих двумя важнейшими свойствами:
-
избирательностью (селективностью) по отношению к определенным ионам;
-
электровозбудимостью, т.е. способностью открываться и закрываться в ответ на изменения мембранного потенциала.
Так же как ионные насосы, ионные каналы образованы макромолекулами белков, пронизывающими липидный бислой мембраны.
Активный и пассивный ионный транспорт. В процессе восстановления после ПД работа калий-натриевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, благодаря чему нарушенное при возбуждении неравенство концентраций Na+ и K+ по обе стороны мембраны восстанавливается. На работу этого механизма тратится около 70 % необходимой клетке энергии.
Таким образом, в живой клетке существует две системы движения ионов через мембрану.
Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии (пассивный ионный транспорт). Он ответствен за возникновение потенциала покоя и ПД и ведет в конечном итоге к выравниванию концентрации ионов по обе стороны клеточной мембраны.
Второй осуществляется против концентрационного градиента. Он состоит в «выкачивании» ионов натрия из цитоплазмы и «нагнетании» ионов калия внутрь клетки. Этот тип ионного транспорта возможен лишь при условии затраты энергии обмена веществ. Его называют активным ионным транспортом. Он ответствен за поддержание постоянства разности концентраций ионов между цитоплазмой и омывающей клетку жидкостью. Активный транспорт - результат работы натриевого насоса, благодаря которому восстанавливается исходная разность ионных концентраций, нарушающаяся при каждой вспышке возбуждения.