

Строение клетки протистов
.pdf4.2. ЦИТОСКЕЛЕТ
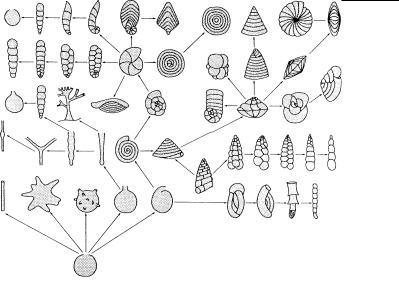
Рис. 4.58. Многообразие форм минерального скелета у протистов. (По: Петрушевская, 1986.)
А – циста акантарии Heteracodon с остатками минеральных игл
(и) внутри, Б–З – кремниевые скелеты полицистин: Б –
Anakrusa, Â – Lithelius nautiloides, Ã – Cladococcus diplospinosus, Ä – Actinomma beroes, Å – Thalassothamnus pinetum, Ж – представитель сем. Pylonidae, З – внутреннее строение Amphisphaera neptunus. и – радиальные иглы, цк – центральная капсула, 1–3 – скелетные оболочки. Масштабные линейки: В – 50 мкм, Г – 75 мкм, Д – 50 мкм, З – 50 мкм.
211

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
форм (рис. 4.58). Обычно это ажурные решетки, формирующие шары (часто вставленные один в другой), конусы, шлемы, короны, цилиндры, башенки и т.д. Все эти скелетные образования иногда называют раковинкой, что неверно, т.к. скелет у полицистин внутренний. Он полностью окружен цитоплазмой и выполняет для клетки скорее опорную функцию, а не защитную. Скелетные элементы исходно формируются внутри цитоп - лазматических матричных пузырьков, или цистерн. При повреждениях скелетных игл (например, отломлен кончик иглы) цитоплазма заполняет поврежденный участок и восстанавл и- вает иглу полностью.
Скелет феодарий образован отдельными иглами, часто полыми внутри. Образование полых игл связано с особенностями их формирования в процессе онтогенеза. Если у полицистин игл ы растут постепенно, заполняя все пространство матричного пузырька, увеличивающегося по мере роста иглы, то у феодарий кремнезем откладывается по периферии пузырька, уже имеющего форму будущей иглы; поэтому внутри сформированной иглы остается полость. Скелет феодарий всегда расположен снаружи от центральной капсулы и состоит из радиальных и тан - гентальных элементов. Отдельные иглы, соединяясь друг с д ругом, образуют ажурные решетки или более плотные структуры , напоминающие раковинки двухстворчатых моллюсков. Скеле т феодарий редко образует геометрически правильные фигур ы и более многообразен, чем у полицистин.
Внутренний неорганический скелет из кремнезема характе - рен также для эбрий и некоторых пединелловых (например, у одной из стадий жизненного цикла Dictyocha) (ðèñ. 4.11).
Весьма многообразны скелетные структуры, построенные на основе кальция. Они встречаются в разных группах протисто в: фораминиферы, гаптофитовые6 (рис. 4.59), инфузории, раковинные амебы, миксомицеты, зеленые водоросли, хризофитовые и динофитовые (Faber, Preisig, 1994). Наиболее широко распространены кальцинированные раковинки среди форами - нифер (рис. 4.60). Обычно они формируют раковинки 3 типов:
6О кальциевых чешуйках кокколитофорид (гаптофитовые) см. в разделе «Покровы»
212

4.2. ЦИТОСКЕЛЕТ
Рис. 4.59. Различные типы чешуек у гаптофитовых водорослей. (По разным авторам.)
1 – Emiliania huxleyi, 2 – лапидолит Laminolithus hellenicus, 3 – калиптолит, 4 – рабдолит Discosphaera tubifer, 5 – зиголит
Corisphaera multipora, 6 – спиральный плаколит Helicosphaera carteri, 7 – плаколит Coccolithus pelagicus, 8 – канеолит Syracosphaera aff. nodosa, 9 – пенталит Braarudosphaera bigelowii.
агглютинированные, составленные из кальцитных игл, и секретируемые раковинки из карбоната кальция с глянцевым вн утренним слоем. Иногда выделяют в особый тип органические р а- ковинки, не имеющие минеральных соединений. Раковинка фораминифер окружена снаружи слоем цитоплазмы, поэтому, строго говоря, их скелет не внешний, а внутренний, и раковин - кой, по-видимому, может называться лишь условно. Раковинки большинства фораминифер многокамерные (имеют от 5 до 30 камер), однако есть также однокамерные и двухкамерные, а у некоторых форм их количество достигает нескольких соте н. В последнем случае камеры могут быть дифференцированы по форме и функции. Стенка раковинки обычно пронизана системой пор и каналов, через которые обеспечивается связь ком -
213
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Рис. 4.60. Многообразие форм раковинок фораминифер и возможные пути их эволюционных преобразований от однока - мерных к многокамерным. (По: Hausmann, Hьlsmann, 1996.)
партментов цитоплазмы клетки между собой. Строение и форма этих структур имеет важное таксономическое значение д ля фораминифер. Снаружи раковинка может быть усилена различной формы выростами и шипами.
Другая группа протистов, имеющих раковинки – это раковинные амебы, или тестации. Многие из них имеют органические кремниевые домики, секретируемые амебой, но немало и других, которые формируют агглютинированные раковинки с минеральными включениями. У филозных амеб рода Cryptodifflugia домик включает кальций в форме карбонатов и фосфатов, которые в виде аморфного вещества покрывают орг а- ническую раковинку снаружи. Органическая раковинка лобозной амебы Trichosphaerium включает кальциевые спикулы, а одна из арцеллинид (Testacealobosia) Paraquadrula секретирует кальциевые пластинки, встраивающиеся в хитиноидный матрикс домика.
214

4.2.ЦИТОСКЕЛЕТ
Óнекоторых инфузорий кальциевые пластинки залегают
âпериферической цитоплазме
под покровами, часто образуя правильные однослойные или двухслойные ряды. Это характерно для представителей родов
Plagiopodon, Coleps, Nolandia. Инфузории из семейства Tintinnidae строят органические домики, которые часто включа- ют кальциевые структуры. Однако они не секретируют их самостоятельно, а используют уже готовые, чаще всего кокколиты.
Депо кальция обнаружено в стенке спорангия некоторых миксомицетов (сем. Didymiidae).
4.2.8.Прикрепительные
органеллы
Многие свободноживущие протисты способны прикрепляться к субстрату и удерживаться на нем при помощи различных образований. Чаще всего сидячие организмы формируют стебелек. Он может быть сократимый, как у сувойки
(рис. 2.43, 4.61), или нет, как у многих других протистов. Его состав и способ образования весьма различаются у разных в и- дов. Это может быть цитоплазматический вырост задней част и клетки как у псевдодендромонад, хоанофлагеллат и некотор ых инфузорий. У некоторых хризофитовых и бикозоецид стебеле к является продолжением домика. Десмоторацидные солнечни - ки формируют стебелек на основе длинной псевдоподии, кото -
215
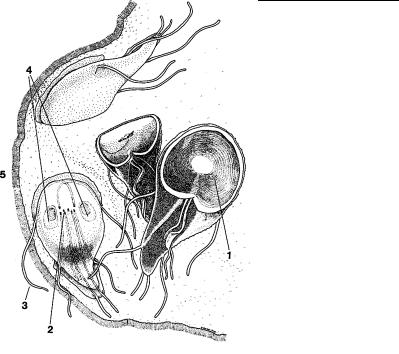
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Рис. 4.62. Внешний вид паразитической дипломонады giardia. Верхняя клетка показана в обычном прикрепленном состоянии к стенке кишечника. (По: Margulis et al., 1993.) 1 – присасывательный диск, 2 – кинетосомы жгутиков, 3 – жгутики, 4 – ядра, 5 – стенка кишечника.
рая служит основой для синтеза внеклеточной трубчатой структуры.
Среди инфузорий встречаются клетки как с цитоплазмати- ческими стебельками, так и со стебельками из внеклеточног о материала.
Необычный стебелек формируют некоторые пединелловые. Например, у Pteridomonas он представлен длинным и тонким цитоплазматическим выростом на заднем конце тела, в который заходит сильно вытянутое микротельце. Когда Pteridomonas отрывается от субстрата и переходит к плаванию, стебелек сохраняется и тянется за клеткой, существенно мешая передви - жению простейшего.
Наиболее своеобразны ножки плодовых тел слизевиков. У клеточных слизей (акразиевые, диктиостелииды) стебельки формируются из отдельных не слившихся, но плотно прилегающих друг к другу клеток. У миксомицетов (неклеточных сли-
216
4.2. ЦИТОСКЕЛЕТ
зевиков) ножка плодового тела образуется из цитоплазмати ческого выроста плазмодия.
Среди других прикрепительных органелл следует отметить различные модификации жгутиков или специальных выросто в клетки. Жгутик часто выполняет функцию сократимого стебе лька (бикозоециды, бодониды) или прикрепляет клетку паразит и- ческого жгутиконосца к стенке кишечника (кинетопластиды ).
Прикрепительные структуры характерны для многих паразитических простейших, обитающих в кишечнике животных, где они постоянно находятся под воздействием пищевого по тока. Для того, чтобы удержаться в определенном участке кише ч- ника, они прикрепляются к его стенке при помощи присосок или крючков. Своеобразные изменения вентральной поверхн о- сти клетки в виде присасывательного диска развиваются у д ипломонады Giardia (рис. 4.62), паразитирующей в кишечнике человека и животных.
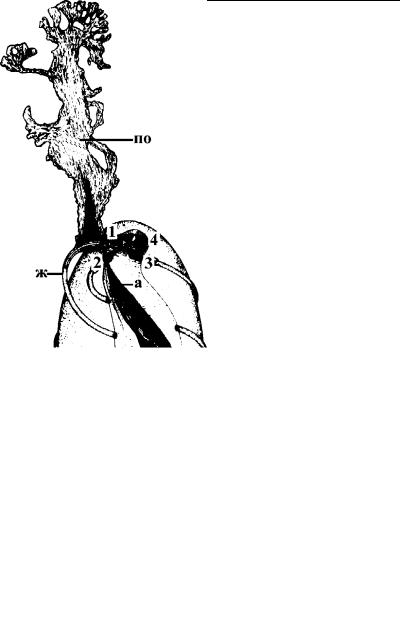
Рис. 4.63. Прикрепительная органелла у оксимонады
Pyrsonympha. (По: Cohrane et al., 1979.) Органелла (по) образована цитоплазматическим выростом, пронизанным фибриллами, которые формируются на основе электронно-плотного материала, окружающего кинетосому и проксимальную часть одного из жгутиков (ж). а – аксостиль, 1–4 – кинетосомы жгутиков.
217

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Мощная присоска в форме прикрепительного диска сложного строения имеется на поверхности клетки паразитичес кой инфузории Trichodina, обитающей в кишке голотурии.
Крючки, или длинные выросты поверхности клетки, которые позволяют простейшему заякориваться в стенке кишечника , развиваются в самых разных группах кишечных паразитов. Так, у паразитических оксимонад из передней части клетки формируется специальная прикрепительная органелла, обр а- зованная цитоплазматическим выростом, который густо про низан микрофиламентами (рис. 4.63). Широкое разнообразие крючков и стилетов демонстрируют эпимериты грегарин из к и- шечника членистоногих (рис. 4.64). Иногда эпимериты формируют тонкие волосовидные отростки, цепляющиеся между эпи -
Рис. 4.64. Эпимериты грегарин, паразитирующих в кишечнике членистоногих. (По: Чорик, Спасский, 1986.) 1 – Gregarina longa, 2 – Sycia inipitata, 3 – Pileocephalus heeri, 4 – Stylocephalus longicollis, 5 – Beloides firnus, 6 – Cometoides crinitus, 7 – Geniorhynchus monnineri, 8 – Echinomera hispida.
218

4.2. ЦИТОСКЕЛЕТ
телиальными клетками (рис. 4.64). Диномонада Haplozoon, паразитирующая в полихетах, имеет сразу 2 приспособления: пу - чок филоподий на переднем конце тела, которые внедряются в поверхность эпителия, и хитиновый подвижный стилет, периодически вонзающийся в поверхность кишечника (Догель, 1951). Описанные случаи дают только примерное представление о многообразии прикрепительных структур клетки протистов .
Многие из них еще совершенно не изучены.
4.2.9. Другие формы движения
Формы движения простейших разнообразны, чему соответствует и многообразие органоидов движения. Как правило, различают 5 основных форм движения - амебоидное, жгутиковое, ресничное, метаболирующее и скользящее. Кроме того, сущес т- вуют и некоторые более специализированные формы двигате льнойактивности.Жгутиковая,ресничная иамебоидная формыдвижения были рассмотрены ранее в связи с описанием цитоскел ета
èпоэтому здесь не приводятся.
Метаболирующее движение
Метаболирующий тип движения был выделен сравнительно
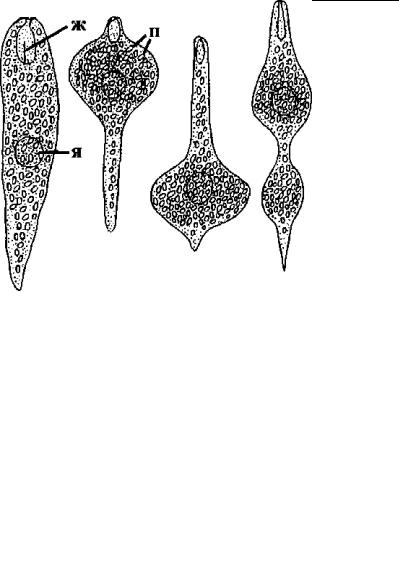
недавно (Серавин, Фролов, 1983), хотя, конечно, давно и хорошо изестен. Некоторые авторы относят его к скользящей фор ме движения. Он характерен для большинства эвгленовых водор ослей, некоторых грегарин и ряда инфузорий и внешне выгляди т как перистальтические или волнообразные изгибания тела клетки (рис. 4.65), за счет которых организм может активно передвигаться. Такая метаболия тела наиболее изучена и хорошо известна преимущественно для эвгленовых (Distigma, Parastasia), поэтому часто называется эвгленоидным движением.
Эвгленовые водоросли способны также к активному изгибанию клетки при повороте в ту или иную сторону. Этот процесс осуществляется благодаря особым покровам эвгленовых – кутикуле, которая состоит из продольных полосок или ребер (см. рис. 4.8). Белковые полосы кутикулы могут скользить относительно друг друга, за счет того, что они ассоциированы с микротрубочками, а те, в свою очередь, связаны с микротру-
219
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
бочками соседних полос при помощи микрофиламентов, которым, вероятно, и принадлежит движущее начало. Во всяком случае, после удаления мембран клетки в результате обработки детергентами вся сократительная система работает в прис утствии ионов Са++ и АТФ. Таким образом, скорее всего, в этом задействована акто-миозиновая система сокращения (Suzaki, Williamson, 1986). Компьютерная модель метаболирующего движения эвгленовых, в которой не происходит изменения дл и- ны полосок, а только их изгибание, вполне удовлетворитель но объясняет его механизм за счет скольжения полосок относи - тельно друг друга (рис. 4.66).
Метаболирующее движение грегарин (рис. 4.67) и некоторых инфузорий определяется другими механизмами. У моноцистид (грегарины) изменение формы тела происходит в результате сокращения фибриллярных структур, поперечными кольцами охватывающих тело клетки по периферии (рис. 4.68). По-видимому, в этом процессе задействованы и микротрубоч-
Рис. 4.65. Изменение формы тела при метаболическом движении эвглениды Parastasia fennica. (По: Вита, Суханова, 1983.) ж – жгутик, п – зерна парамилона, я – ядро.
220