

Строение клетки протистов
.pdf4.2. ЦИТОСКЕЛЕТ
Рис. 4.44. Полиподиальная (1) и моноподиальная (2,3) формы голых амеб (По: Смирнов, Гудков, 2000.)
1 – Amoeba, Chaos; 2 – Amoeba, Chaos; 3 – Saccamoeba, Hartmannella, Trichamoeba.
образование псевдоподий при изменении направления движения простейшего.
Филоподии. Для многих амебоидных протистов характерны филоподиальные выросты – тонкие длинные иногда ветвящиеся, не анастомозирующие между собой псевдоподии. Они характерны для филозных амеб и служат как для движения, так и для питания. Их внутренний скелет состоит из актиновых микрофиламентов. Микротрубочки в них почти не встречаются. Лишь в некоторых филоподиях обнаруживаются одиноч- ные микротрубочки. Филоподии способны медленно сокращаться (втягиваться, вытягиваться, изгибаться).
По внешнему виду клеток у филозных амеб (как, впрочем, и у многих других амеб) довольно четко выделяются две «жизне нные формы»: субстратная, или «распластанная», которая характе рна для лежащих на субстрате амеб, и флотирующая, или «радиаль - ная», характерная для парящих в толще воды амеб (рис. 4.45). В первом случае клетка уплощена и лежит на субстрате, актив но перемещаясь. Во втором случае клетка имеет шаровидную форму с радиально расходящимися филоподиями, пассивно парит в то л- ще воды и перемещается с ее потоками. Представители одних видов могут принимать обе формы, а для других описана только флотирующая форма или только субстратная.
191
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
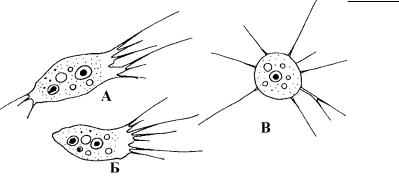
Рис. 4.45. Жизненные формы филозной амебы Nuclearia simplex (Ïî: Pernin, 1976.)
А – субстратная форма с двумя пучками псевдоподий, Б – субстратная форма с одним пучком псевдоподий, В – флотирующая форма.
Данные о строении и функционировании псевдоподий у филозных амеб отрывочны, а механизм функционирования филоподий никогда не был объектом пристального внимания. Пе - редвижение клетки осуществляется медленно. Вся клетка ползет как единое целое необъяснимым образом, а передние филоподии вытянуты по направлению движения, как бы «ощупывая» субстрат (рис. 4.45). Лишь иногда отдельные филоподии прикрепляются к субстрату дистальными кончиками.
В литературе описано перемещение клетки при помощи прикрепления дистальной части филоподии к субстрату и дальн ейшего «подтягивания» клетки путем сокращения филоподии. О д- нако более поздние наблюдения не подтверждают этих описаний.
Механизм сокращения этих псевдоподий почти не изучен. Для флотирующей формы Nuclearia было показано, что сокращение филоподии происходит за счет быстрого формирования эндо цитозных пузырьков, которые отшнуровываются внутрь клетки и транспортируются в ее цитоплазму (Mignot, Savoie, 1979). Таким образом, в амебе формируется запас мембран, которые потребуются ей в дальнейшем для формирования новых длинных филоподий.
192

4.2. ЦИТОСКЕЛЕТ
Рис. 4.46. Внешний вид ретикулоподиальной амебы
Reticulomyxa filosa
(По: Bovee, 1985.) р – ретикулоподии.
У многих филозных амеб, имеющих раковинку, часть гиалоплазмы выходит во время движения через устье раковинки и распластывается по субстрату. Образуется гиалиновый дис к неправильной формы, от которого берут начало филоподии.
Ретикулоподии (ранее их называли ризоподиями) - тонкие ветвящиеся и анастомозирующие цитоплазматические вырос - ты (рис. 4.46). Они служат для прикрепления клетки к субстрату, ее перемещения, а также для захвата пищи. Перемещение клетки происходит довольно медленно, путем «прилипания» ре-
193

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
тикулоподий к субстрату и дальнейшего подтягивания клет ки в результате их сокращения. Неподвижные клетки формируют из ретикулоподий настоящую ловчую сеть, в которую легко п о- падаются различные мелкие организмы.
В отличие от филоподий, внутри ретикулоподий всегда обнаруживаются микротрубочки. В зависимости от толщины ретикулоподий, микротрубочки могут быть одиночными или фор - мировать толстые пучки. Однако во всех описанных случаях микротрубочки не связаны между собой и не образуют упорядоченных структур, которые мы видим в аксоподиях и жгутиках. В крупных ретикулоподиях хорошо заметно движение пищевых и других частиц цитоплазмы, которое происходит, повидимому, при участии микротрубочек. У фораминифер, например, при описании этих процессов используются термины «реоплазма» и «стереоплазма», которые применяются и в отношении аксоподий (см. стр. 61–64,93).
Ретикулоподии часто содержат небольшие заметные в световой микроскоп гранулы. Такие псевдоподии часто называют гранулоретикулоподиями, а их обладателей - гранулоретику - лозными амебами. До недавнего времени существовал таксон Granuloreticolosea, объединявший все подобные организмы. Ретикулоподиальные формы встречаются в разных таксонах протистов как среди простейших (фораминиферы), так и среди водорослей (хлорарахниды).
По-видимому, ретикулоподии характерны и для акантарий, хотя обычно их называют филоподиями (Решетняк, 1981). Эти псевдоподиальные выросты находятся на поверхности экто п- лазматического кортекса клетки, и представлены тонкими в етвящимися и анастомозирующими между собой псевдоподиями, формирующими сеть для захвата пищи. Внутреннее строение этих псевдоподий не изучено, но по внешнему виду их, вероятно, следует считать ретикулоподиями.
Морфотипы
Форма тела живой амебы непрерывно изменяется, поэтому детальное и точное описание его, как это принято для други х протистов, практически невозможно. Однако еще в 1926 году А.А.Шаффер (Schaeffer, 1926) обнаружил, что при активном,
194

4.2. ЦИТОСКЕЛЕТ
направленном перемещении амеба принимает характерный именно для данного вида облик, основные черты которого со - храняются все время, пока организм продолжает движение. Другими словами, во время движения (локомоции) амеба принимает динамически стабильную форму, которая в каждый пос - ледующий момент немного отличается от предыдущего, но в целом сохраняет свои характерные черты все время, пока клет ка направленно перемещается. Поэтому при изучении этих протистов исследователи невольно сравнивают между собой им енно локомоторные формы амеб.
К настоящему времени описано уже несколько сотен видов голых амеб, каждый из которых обладает своими особенностя - ми организации локомоторной формы. И хотя они плохо поддаются формальному описанию, в последнее время были предприняты попытки найти несколько основных «образов» движущихся амеб, которые были названы «морфотипами» (Смирнов, Гудков, 2000). По определению авторов, морфотип – это обобщенный образ локомоторной формы амебы, включающий всю совокупность признаков, описывающих ее динами- чески стабильную организацию. Например, полиподиальная и моноподиальная формы амеб (рис. 4.44) представляют собой 2 разных морфотипа. Разработка концепции морфотипов в настоящее время еще не завершена, поэтому полная классифика - ция их не приводится.
4.2.5. Цитоскелет амеб и амебоидное движение
Эктоплазма амеб пронизана сеточкой микрофиламентов из актина и миозина, которая, собственно, и представляет собо й скелет клетки. Этот кортикальный слой, или корсет из микро - филаментов, связан с плазмалеммой амебы и полностью окружает все содержимое клетки, даже в тех местах, где эктоплаз ма почти незаметна. Корсет из микрофиламентов обладает морф о- логической и физиологической полярностью. Причем актино - вые и миозиновые филаменты распределяются вдоль клетки п о- разному. У движущейся амебы актин образует довольно тонки й слой на переднем конце клетки, в средней части его толщина
195

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
увеличивается, а на заднем конце (в районе уроида) опять уменьшается (рис. 4.47). Миозиновые волокна также образуют тонкий слой на переднем конце клетки, в средней части ег о толщина увеличивается, а в задней достигает максимальной толщины. Различается и ориентация волокон цитоскелета. В передней трети тела движущейся амебы актиновые филаменты ориентированы вдоль поверхности тела. Их продольные ряды связаны поперечными мостиками как между собой, так и с пла з- малеммой. В задней части тела актиновые филаменты образуют сложную трехмерную сеть, в которой залегают толстые ми о- зиновые филаменты. Эти данные по строению цитоскелета аме б из семейства Amoebidae (Amoeba proteus è Chaos carolinense), а также амебы Dictyostelium были получены сравнительно недавно. Они послужили основой для гипотезы, объясняющей амебоидное движение у амеб. Так называемая «теория генерализованного кортикального сокращения», предложеная Гребецким (Grebecki, 1982), объясняет амебоидное движение следующим образом.
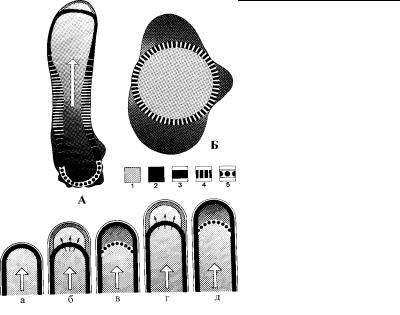
Трехмерное сокращение акто-миозинового кортекса приводит к сжатию эндоплазмы, в результате чего она направляется к переднему концу амебы, где кортекс наиболее тонкий. Сюда же приносятся молекулы глобулярного актина (G-актина), которые постоянно образуются на заднем конце клетки в резул ь- тате деполимеризации актиновых микрофиламентов (F-акти- на), входящих в состав кортекса.
В результате этого сокращения кортекса в эндоплазме создается повышенное давление, которое продавливает цитоплазму клетки сквозь слой микрофиламентов на ее переднем конце к ак сквозь сито. В результате плазмалемма переднего конца амебы отслаивается от корсета из микрофиламентов и выпячивается наружу. Молекулы G-актина (в отличие от крупных включе- ний эндоплазмы) также проходят сквозь него и попадают в пр о- странство между цитоскелетом и плазматической мембрано й в растущую лобоподию (рис. 4.47). На внутренней поверхности плазмалеммы расположены специальные центры, которые вызывают полимеризацию актина, т.е. G-актин превращается в F-актин формируя при этом новый кортикальный слой микро-
196
4.2. ЦИТОСКЕЛЕТ
Рис. 4.47. Схема строения и преобразования цитоскелета амебы во время движения. (По: Stockem, Klopocka, 1988.)
А – цитоскелет амебы при нормальном движении, Б – цитоскел ет амебы в состоянии покоя. 1 – цитоплазма в состоянии золя, 2 – цитопл азма в состоянии геля, 3 – только что сформированный филаментозный слой, 4 – сократившийся филаментозный слой, 5 – разрушенный слой филаментов. а–д – процесс формирования псевдоподии у движ ущейся амебы в соответствии с гипотезой генерализованного корт икального сокращения. Большая стрелка показывает направление движ ения эндоплазмы, маленькие стрелки указывают прохождение цит оплазмы с молекулами G-актина сквозь корсет из микрофиламентов.
филаментов, связанных с плазматической мембраной. Вновь сформированный слой микрофиламентов начинает сокращать - ся и оказывает на цитоплазму давление, направленное назад . Вследствие этого рост лобоподии прекращается. В это же вр емя происходит деполимеризация отслоившегося от плазмалемм ы участка кортекса.
В дальнейшем, если движение амебы продолжается в том же направлении, все этапы превращения G-актина в F-актин и деполимеризации последнего повторяются: новый слой актино -
197
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
вых филаментов отслаивается от мембраны, продолжая при этом сокращаться, движется назад, тогда как плазмалемма под напором фильтрующейся сквозь него эндоплазмы движется в противоположную сторону. Это вызывает дальнейший рост гиалиновой шапочки и, соответственно, лидирующей псевдоподии (рис. 4.47).
|
Изложенная гипотеза хоро- |
|
|
шо объясняет многие аспекты |
|
|
локомоции амеб, однако только |
|
|
тех, которые похожи на Amoeba |
|
|
proteus. Она объясняет и тот |
|
|
факт, что во время движения |
|
|
амеба периодически прикреп- |
|
|
ляется к субстрату и открепля- |
|
|
åòñÿ îò íåãî (ðèñ. 4.48). Îäíà- |
|
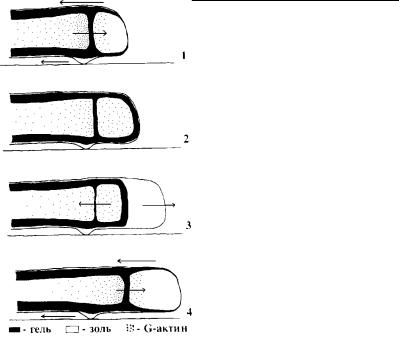
Рис. 4.48. Реорганизация |
ко даже среди лобозных амеб |
|
существует множество видов с |
||
системы микрофиламентов на |
||
другими типами движения, |
||
переднем конце растущей |
||
лобоподии. Вид сбоку. |
которые невозможно объяс- |
|
(Ïî: Grebecki, 1990.) |
нить предложенной схемой. |
|
1–4 – последовательные стадии |
||
В качестве исторической |
||
изменения организации |
||
микрофиламентов. Стрелки |
справки можно привести пред- |
|
показывают направление |
шествующие гипотезы амебо- |
|
движения тока цитоплазмы и |
||
идного движения. Теория Ма- |
||
плазмалеммы лобоподии. |
||
Видно перемещение клетки |
ста (Mast, 1926), или «теория |
|
вправо по отношению к точке |
потока под давлением» заклю- |
|
прикрепления к субстрату. |
||
чается в том, что в результате |
||
|
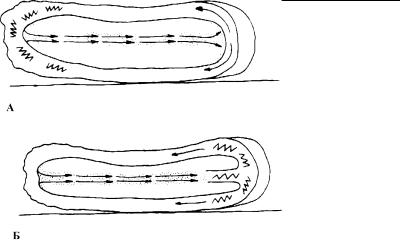
сокращения эктоплазмы на заднем конце клетки создается и з- быточное давление, которое вызывает движение эндоплазмы в передний конец клетки (рис. 4.49). Достигая гиалиновой шапочки, золеобразная эндоплазма растекается во все сторон ы псевдоподии наподобие струй фонтана (так называемое «фон -
198
4.2. ЦИТОСКЕЛЕТ
Рис. 4.49. Схематическое изображение механизмов амебоидного движения по Масту (А) и Аллену (Б).
Стрелки показывают направление движения цитоплазмы, а зигзагообразные линии – зону сокращения цитоплазмы.
танирующее движение эндоплазмы»). В кортикальной зоне клетки эта эндоплазма превращается в эктоплазму, имеющую гелеобразное состояние. В то же время на заднем конце амебы происходит обратное превращение эктоплазмы в эндоплазм у. Эти процессы происходят так быстро, что создается впечатл е- ние плавного непрерывного тока цитоплазмы, в результате чего клетка продвигается вперед.
Аллен (Allen, 1961) считал, что зона сокращения эктоплазмы находится не на заднем, а на переднем конце амебы (рис. 4.49). При этом золеобразная эндоплазма на переднем конце тела а мебы переходит в гелеобразное состояние. В результате новая порция эндоплазмы как бы подтягивается к переднему концу особи. Движение цитоплазмы в клетке идет по той же схеме, что и в гипо тезе Маста. Эктоплазма переходит в состояние золя в зоне уроид а.
Оба автора опирались на детальные наблюдения за движущейся амебой (преимущественно Amoeba proteus) и экспериментальные воздействия на нее, предлагая единый механизм, вызывающий амебоидное движение. Однако эти гипотезы
199

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
объясняли лишь некоторые аспекты амебоидного движения. Формы движения амеб действительно весьма разнообразны. Например, следует четко различать амебоидную активность , которая приводит к образованию псевдоподий у неподвижно й амебы и позволяет ей формировать пищевые псевдоподии для захвата пищи, и собственно амебоидное движение, которое н е всегда связано с изменением формы тела. Поэтому естествен - ным выглядит предположение Л.Н.Серавина о существовании множественных механизмов, обеспечивающих амебоидное дв и- жение (Серавин, 1967). Суть этой гипотезы заключается в том, что у всех амебоидных клеток имеется одинаковый набор из нескольких механизмов, которые обеспечивают все разнооб разие амебоидных клеток и форм их движения. Собственно же различия обуславливаются степенью участия того или иног о механизма в двигательной активности амебоидного организм а. В свете этих представлений гипотеза генерализованного со кращения представляет собой описание лишь одного из возможных механизмов амебоидного движения.
4.2.6. Другие скелетные образования
Многообразие скелетных структур протистов исключительно велико, и приведенные выше примеры далеко не исчерпывают его. Существует много других структур, которые менее часто встречаются у протистов, а некоторые из них уникаль ны. В этой главе будут рассмотрены другие скелетные образова ния клетки протистов, а также ее различные выросты и другие ос о- бенности.
Аксостиль
Аксостиль обычно представлен группой плотно упакованны х микротрубочек, идущих вдоль продольной оси клетки (рис. 4.30) , и действительно составляет внутренний осевой скелет мно гих парабазалий и оксимонад. У многих видов аксостиль обладае т подвижностью: он может изгибаться, скручиваться, поворачи - ваться вокруг своей продольной оси, инициируя изменение формы клетки и даже ее движение. На поперечном срезе он выг - лядит в виде пучка или свернутой продольно широкой ленты
200