

Строение клетки протистов
.pdf4.1. ПОКРОВЫ
Рис. 4.10. Схема строения типичной пелликулы инфузорий (А), кутикулы у сосущих инфузорий (Б) и энтодиниоморф (В).
ал – альвеола, мт – микротрубочки, пл – плазмалемма, фп – филаментозный пласт, эк – эктоплазма, эп – эпиплазма.
(А – по: Dodge, 1973, Б, В – по: Серавин, Герасимова, 1979.)
плазмалемму плоских пузырьков – альвеол. Альвеолы плотно смыкаются краями друг с другом, располагаясь в один слой (рис. 4.1 м; 4.10 А). Такие покровы называются пелликулой и характерны для альвеолат (инфузорий, споровиков и динофи - товых).
Как у инфузорий, так и у динофитовых можно проследить все переходы, отражающие, по-видимому, этапы становления пелликулы, от рыхло расположенных под плазмалеммой альвеол, которые не смыкаются краями друг с другом и более похожи на обычные пузырьки, чем на уплощенные цистерны (такие покровы иногда называются пропелликулой), до настоящ ей пелликулы. Альвеолы динофитовых часто содержат различны е структуры – от тонких аморфных образований неизвестной п рироды до плотных целлюлозных пластинок. В последнем случае такие усиленные покровы называют текой, или амфиесмой (рис. 4.1 н). У зоитов споровиков количество альвеол невелико, поэтому их немногочисленные контакты друг с другом бы - вает очень трудно обнаружить. Кроме того, эти альвеолы на-
141

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
столько уплощены, что выглядят на срезах как две почти при - легающие друг к другу мембраны, которые принято называть внутренним мембранным комплексом. Между тем и по происхождению и по строению покровы споровиков являются пелликулой.
На основе пелликулы формируется и панцырь диатомовых водорослей. Электронно-плотные кремниевые структуры зак - ладываются внутри подстилающих плазмалемму альвеол, которые затем сливаются друг с другом по мере формирования панцыря. Следует заметить, что под альвеолами пелликулы по- чти всегда обнаруживаются микротрубочки.
Нередко под пелликулой инфузорий залегают мощные пуч- ки микротрубочек, идущие в продольном и поперечном направ - лениях (рис. 4.10 В). Они могут дополнительно усиливаться слоем плотной эктоплазмы (эпиплазмы) или фибриллярными структурами (рис. 4.10 Б), формируя кутикулу.
В редких случаях под плазмалеммой формируется плотно прилегающая к ней дополнительная мембрана (рис. 4.1 л). На поперечных срезах такие покровы выглядят состоящими из д вух мембран4 . Происхождение и природа внутренней мембраны неизвестны. Встречаются такие покровы крайне редко и обнару - жены как постоянные структуры пока только у апузомонад. П о- хожее образование имеется у некоторых представителей гемимастигофор (Hemimastix). Правда, оно выглядит как тонкий фибриллярный слой, который авторы называют уплотнением эпиплазмы (Foissner et al., 1988).
Эпиплазматический слой встречается в покровах разных пр о- тистов и может оказаться весьма важным для установления ф и- логенетических связей между их группами (Philippe, Adoutte, 1998). Он образован белками, которые, как оказалось, близки по своим иммунологическим свойствам у цилиат, динофитовых и эвгленовых. Кроме того, последовательности нуклеотидов ге-
4Ранее (Карпов, 1986, 1990) я выделял этот тип особо, называя его «Формирующимся за счет изменения самой плазмалеммы». Одн ако изменения плазмалеммы здесь, фактически, нет. К ней лишь прилегает внутренняя мембрана, которая имеет такую же толщину и такое же трехсл ойное строение, как и наружная плазматическая мембрана.
142
4.1. ПОКРОВЫ
нов, кодирующих эти белки у Euglena (Marrs, Bouck, 1992) и инфузории Pseudomicrothorax (Huttenlauch et al., 1995), демонстрируют поразительное подобие, что свидетельствует о вы сокой степени гомологии этих белков.
Таблица 1. Типы покровов у протистов
Тип покровов |
|
|
клеточнаястенка |
|
|
|
|
|
двойнаямембрана |
Таксон |
гликостили |
чешуйки |
домик или панцирь |
тубулемма |
пелликула и ее производные |
кутикула |
перипласт |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Microsporidia |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Chytridiomycota |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Myxozoa |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Chlorophyta |
- |
+ |
+ |
- |
- |
- |
- |
- |
- |
Rhodophyta |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Glaucophyta |
- |
- |
+ |
- |
- |
+ |
- |
- |
- |
Cryptophyta |
- |
+ |
- |
- |
- |
- |
- |
+ |
- |
Chrysophyceae |
- |
+ |
+ |
+ |
- |
- |
- |
- |
- |
Bicosoecida |
- |
+ |
- |
+ |
- |
- |
- |
- |
- |
Eustigmatophyceae |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Phaeophyceae |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Bacillariophyceae |
- |
- |
- |
+ |
- |
+ |
- |
- |
- |
Haptophyta |
- |
+ |
+ |
- |
- |
- |
- |
- |
- |
Oomycetes |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Labyrinthomorpha |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
Opalinata |
- |
- |
- |
- |
ã |
- |
- |
- |
- |
Hyphochytridea |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Euglenoidea |
- |
- |
- |
+ |
- |
- |
+ |
- |
- |
Kinetoplastidea |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
Choanomonada |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
Retortamonadida |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
Dinophyta |
- |
+ |
+ |
- |
- |
+ |
- |
- |
- |
Apicomplexa |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
Ciliophora |
- |
- |
- |
+ |
- |
+ |
+ |
- |
- |
Plasmodiophora |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Mycetozoa |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
Ризоподы |
+ |
+ |
- |
+ |
- |
- |
- |
- |
- |
Foraminifera |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
Heliozoa |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
Polycystinea |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
Apusomonadida |
- |
- |
- |
- |
- |
- |
- |
- |
+ |
Hemimastigida |
- |
- |
- |
- |
- |
- |
+ |
- |
- |
Thaumatomonadida |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
Обозначения: “-“ - отсутствует, “+” - имеется, г - гребенчатая тубулемма.
143
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
4.2.Цитоскелет
Âоснове скелетных образований цитоплазмы лежат два типа структур – микрофиламенты и микротрубочки. Их роль в жизни клетки протиста чрезвычайно существенна и многооб разна. Микрофиламенты имеют диаметр 4–10 нм. Они могут быть образованы разными белками – актином, миозином, центрином, ассамблином, динеином, нексином, спазмином и многими другими еще не идентифицированными белками. Микрофиламенты обычно формируют как сократимые, так и несократимые фи б- риллярные тяжи, которые могут быть связаны с кинетосомами , аксонемами, спикулами, ядром, хлоропластами, митохондриями, плазмалеммой. Корсет из микрофиламентов у амеб обеспе - чивает движение этих простейших. Микрофиламенты участву - ют в питании и работе сократительной вакуоли. Довольно ча сто они ассоциированы с микротрубочками, обеспечивая движен ие частиц цитоплазмы внутри клетки.
Микротрубочки обычно имеют диаметр 24–26 нм, хотя встре-
чаются и более толстые – до 40 нм. Они состоят, как правило, из глобулярных белков αα- è ββ-тубулина, которые присутствуют в цитоплазме в виде димера – двух связанных между собой
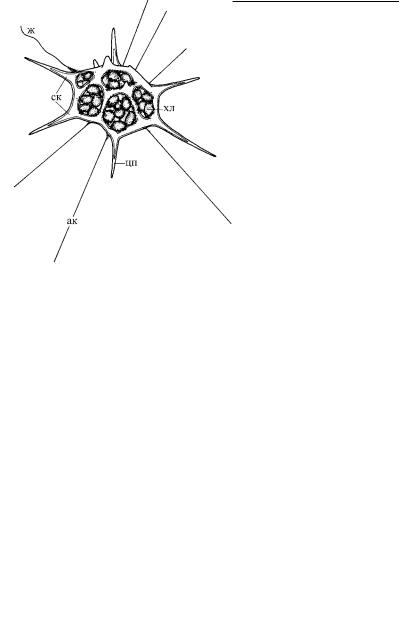
Рис. 4.11. Скелетные образования
Dictyocha. (Ïî: Moestrup, Thomsen, 1990.)
ак – аксоподия, ж – жгутик, ск – скелет, хл – хлоропласт, цп – цитоплазма, обволакивающая цитоскелет.
144
4.2. ЦИТОСКЕЛЕТ
глобул. Микротрубочки формируются из димеров тубулина в результате самосборки. Микротрубочки выполняют в клетке как опорную, так и сократительную функции. Они располагаются под покровами многих инфузорий и жгутиконосцев, определяя форму клетки, входят в состав ресничного и жгутикового аппарата, аксоподий, ретикулоподий, сократительной вакуоли и прочих структур. За счет микротрубочек, содержащих и χχ-тубулин, в процессе деления ядра формируется митотическое, или ахроматиновое, веретено, обеспе- чивающее расхождение хромосом в анафазе. Микротрубочки участвуют в формировании перегородки между до- черними клетками делящейся водоросли, компартментализации цитоплазмы
у крупных инфузорий, в транспорте как мелких, так и крупных вакуолей внутри клетки и выполняют многие другие функции.
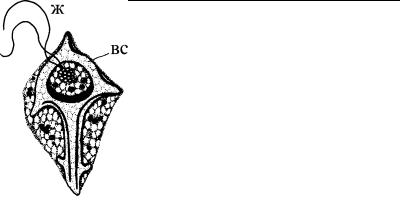
У некоторых протистов (акантарии, феодарии, солнечники) внутренний скелет клетки усиливается за счет минеральны х игл, состоящих из солей стронция или кремния. Мощный и монолитный внутренний скелет найден на одной из стадий жизнен - ного цикла силикофлагеллаты Dictyocha (Pedinellidea) (рис. 4.11), а также у некоторых нетипичных динофитовых (эбрии) (рис. 4.12).
Мощная раковинка некоторых полицистин также может от- части считаться внутренним скелетом (рис. 2.54), т.к. их цитоплазма часто обволакивает раковинку снаружи. Возможно , функция минерального скелета заключается не столько в по д- держании формы клетки, сколько в обеспечении парения план - ктонных организмов в толще воды и передвижения.
В следующей главе будет рассмотрена одна их самых важных частей цитоскелета протистов – система жгутика/реснички.
145

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
4.2.1. Жгутиковый (ресничный) аппарат
Подвижность очень многих протистов осуществляется при помощи жгутиков или ресничек. Те и другие устроены одинаково, но исторически термин «реснички» появился раньше дл я обозначения многочисленных подвижных выростов (ундулиподий) на поверхности клеток инфузорий. Конечно, формально правильнее называть все эти структуры ресничками, одна ко тот и другой термины широко встречаются в литературе, поэ тому разумно использовать их оба. Традиционно считается, что реснички есть у инфузорий, а у других протистов – жгутики. Реснички обычно многочисленны и короче жгутиков, но, с другой стороны, сравнительно короткие жгутики опалин часто н а- зывают ресничками, т.к. они многочисленны, и, в то же время, не принято называть ресничками многочисленные жгутики г и- пермастигин.
У большинства протистов количество жгутиков невелико (от 1 до 8 на клетку), хотя есть немало и многожгутиковых форм. Принято различать монады по внешнему виду жгутиков, их ко - личеству и способу биения (рис. 4.13). Жгутики называются гомодинамными, если их движение сходно и согласованно, и гетеродинамными, если они имеют разные типы биения.
Наиболее часто выделяют 4 группы жгутиконосцев. 1) Изоконтные формы имеют одинаковые гомодинамные жгутики. К ним относится большинство подвижных клеток зеленых водо - рослей. 2) Анизоконты имеют жгутики разной длины, но они также не отличаются по внешнему виду и способу биения. Такие жгутики широко распространены среди бесцветных жгутиконосцев и водорослей. 3) Гетероконтные формы имеют 2 раз - ных по внешнему виду и расположению гетеродинамных жгутика. Они характерны для подвижных клеток водорослей, содержащих хлорофилл c, зооспоровых грибов и многих бесцветных жгутиконосцев. Двигательный жгутик гетероконтн ых форм направлен вперед и обычно опушен мастигонемами (см. стр. 151). Второй жгутик (рекуррентный, или рулевой) направлен назад, лишен мастигонем и обычно пассивен, или обладае т иным характером движения. По этой классификации, гетеро-
146

4.2. ЦИТОСКЕЛЕТ
Рис. 4.13. Варианты расположения жгутиков у протистов. (По: Саут, Уиттик, 1990.)
А – изоконтная форма, Б, Ж – гетероконтные жгутиконосцы с одним билатерально опушенным жгутиком, В – изоконтный жгутиконосец с гаптонемой (средний вырост), Г, Д – изоконтные монады с 4 жгутиками (на рис. Д они выходят из жгутикового кармана), Е – стефаноконтный жгутиконосец, З – гетероконтный жгутиконосец с поперечным и продольным жгутиками, И–Л – одножгутиковые протисты с билатерально (И), унилатерально (К) опушенным жгутиком и с гладким жгутиком (Л).
контными следует считать монады динофитовых и эвгленовы х, несмотря на то, что оба их жгутика опушены мастигонемами, правда, не трубчатыми, а простыми. 4) Стефаноконтные формы имеют венчик из 30–40 жгутиков на переднем конце клетки. К ним относятся многожгутиковые гаметы и зооспоры зеленых водорослей порядка Oedogoniales.
Одножгутиковые формы обычно не выделяются в особую группу. Многие из них справедливо рассматриваются как утратившие второй жгутик особи, т.к. у подавляющего большинства наряду со жгутиковой есть и безжгутиковая кинетосом а. За пределами этой классификации остаются многожгутиков ые
147

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
формы (опалины, гемимастигофоры, зооспоры некоторых хитридиевых грибов, Stephanopogon) и все инфузории.
В эпоху световой оптики исследователи часто придавали таксономический смысл этим группировкам, основанным на особенностях взаимного расположения жгутиков. Например , зеленые водоросли называли изоконтами, желто-зеленые – ге - тероконтами. Хотя, конечно же, эти группы жгутиконосцев представляли лишь определенные морфотипы. По мере того, как одинаковые морфотипы жгутиковых клеток стали обнаруживать в разных таксонах (например, гетероконты встре- чаются и среди зеленых флагеллат), от этой традиции отказа - лись. Однако термин «гетероконты» (или «страминопилы») довольно часто применяется в настоящее время для большой группы протистов, гетероконтные клетки которых имеют на переднем жгутике трубчатые мастигонемы. Таким образом, термин «гетероконты» оказывается эже его первоначального значения. Он не включает гетероконтные клетки динофитовых (для них иногда употребляют термин «диноконтный») и бодонид (кинетопластиды).
Независимо от расположения жгутиков/ресничек на теле клетки, внешняя их часть – ундулиподия – устроена весьма о д- нообразно. Кроме ундулиподии, в составе жгутикового аппарата выделяют переходную зону, кинетосому (базальное тел о)
èкорешковую систему (рис. 4.14).
Ундулиподия и ее поверхностные элементы
Ундулиподия обычно имеет одинаковую толщину по всей
длине (около 200–250 нм). Скелет ундулиподии образован аксонемой, состоящей из 9 пар периферических и 2 центральных микротрубочек, и ассоциированных с ними других структур. Кончик жгутика может быть тупым (периферические и центральные трубочки аксонемы одинаковой длины) или вытянутым в тонкий бичевидный отросток – акронему, которая обыч - но образована парой центральных микротрубочек, «обтянут ых» плазмалеммой.
Почти у всех свободноживущих жгутиконосцев на двигатель - ном жгутике есть чешуйки или мастигонемы, а у некоторых криптомонад и динофитовых жгутики покрыты и мастигоне-
148

4.2. ЦИТОСКЕЛЕТ
Рис. 4.14. Обобщенная схема строения жгутикового аппарата. (Ориг.)
ак – аксосома, бк – безжгутиковая кинетосома, вм – вторичны е микротрубочки, ж – жгутик, жк – жгутиковая кинетосома, ма – трехчленные трубчатые мастигонемы, ч – жгутиковые чешуйк и, пм – периферические дублеты микротрубочек аксонемы, пп – поперечная пластинка, пс – переходная спираль, пф – перехо д- ные фибриллы, р –фибриллярный корешок системы II (ризопласт), тм – триплеты микротрубочек кинетосомы, фс – фибриллярный мостик между кинетосомами, цм – центральные микротрубочки аксонемы, я –ядро, r1 – ребристый корешок с отходящими вторичными микротрубочками (вм), r2–r4 – микротрубочковые корешки.
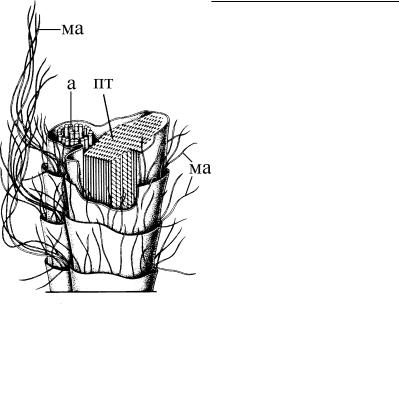
мами, и чешуйками. Принято различать простые мастигонемы и трубчатые. Простые мастигонемы представляют собой тонк ие волоски, или нити, отходящие от поверхности жгутика. Наиболее характерны они для эвгленовых, кинетопластид и динофитовых (табл. 2). На жгутиках эвгленовых есть длинные (до 5 мкм) и короткие (0,5 мкм) волоски (рис. 4.15). Длинные крепятся у них, как и у кинетопластид, с одной стороны жгути - ка, тогда как короткие по спирали опоясывают всю поверхно сть
149
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
|
жгутика. Значительно реже |
||
|
встречаются мастигонемы у |
||
|
хлорофитовых и гаптофи- |
||
|
товых (табл. 2). Их простые |
||
|
волоски также различают- |
||
|
ся по строению и располо- |
||
|
жению на жгутиках, но |
||
|
таких данных немного, по- |
||
|
этому их трудно системати- |
||
|
зировать. |
|
|
|
Трубчатые мастигонемы |
||
|
могут быть двух- и трех- |
||
|
членные. Большой сложно- |
||
|
сти достигают строение и ор- |
||
|
ганизация |
трубчатых |
|
|
трехчленных мастигонем на |
||
|
двигательном жгутике стра- |
||
|
минопилов (рис. 4.16). Они |
||
Рис. 4.15. Схема организации |
имеют, по-видимому, гли- |
||
жгутика эвгленовой водоросли |
копротеиновую природу, и |
||
Anisonema. (Ïî: Mignot, 1966.) à – |
состоят из короткой базаль- |
||
аксонема, пт – параксиальный тяж. |
|||
ной части (0,2–0,3 мкм), |
|||
На поверхности жгутика показаны |
|||
длинные и короткие простые |
длинного полого стержня |
||
мастигонемы (ма). |
(0,7–0,8 мкм) и дистальной |
||
|
|||
части (до 0,5 мкм), образованной терминальными филаментами, количество которых варьирует. Например, у золотистых водорослей (Chrysophyceae) их обычно 3, один из которых длиннее остальных, у эустигматофитовых всего 1 филамент, у зооспор траустохитридиевых 2 неравных филамента.
Мастигонемы празинофитовых образованы полисахаридами. Их строение оказалось более сложным, чем предполагалось ранее (рис. 4.17). Почти у всех мастигонем обнаружено четырех- членное строение: проксимальный, или заякоривающий, филамент, толстая трубчатая часть, дистальная часть из мелких субъединиц и тонкий терминальный филамент. Между трубча- той частью и дистальными субъединицами часто появляется промежуточная часть, а терминальный филамент у таких мас-
150