

Строение клетки протистов
.pdf4.2. ЦИТОСКЕЛЕТ
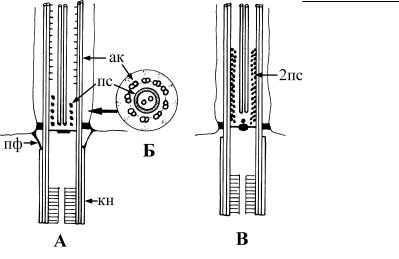
Рис. 4.21. Строение переходной зоны жгутика у зеленых водорослей на продольном (А) и поперечных (Б) срезах. (По: Grain et al., 1988.) Большие стрелки указывают места соответствующих поперечных срезов.
ак – аксонема, зв – звездчатая структура, кн – кинетосома, п ф – переходные фибриллы.
роко используется для определения точного положения отх о- дящих от кинетосомы корешков при 3-мерной реконструкции жгутикового аппарата по ультратонким срезам.
Длина кинетосомы варьирует у разных видов от 50 до 1300 нм. Большинство мастигофор имеет 2 жгутика и 2 соответствующие кинетосомы. Даже у инфузорий кинетосомы в рядах (кинетах)
преимущественно собраны попарно. У одножгутиковых видов тоже, как правило, 2 кинетосомы, одна из которых безжгутиковая (рис. 4.14). Очевидно, что двужгутиковое состояние является наиболее широко распространенным и, по-видимому, эволюционно консервативно. Одножгутиковое состояние мо над принято объяснять редукцией второго жгутика, которая в кр айнем варианте может приводить к исчезновению и кинетосомы . Те организмы, которые имеют только один жгутик и одну кинетосому, принято называть истинно одножгутиковыми. В на-
161
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Рис. 4.22. Строение переходной зоны жгутика у хризофитовых на продольном (А) и поперечном (Б) срезах, и у зооспор оомицетов на продольном срезе (В). (По: Grain et al., 1988.) ак – аксонема, кн – кинетосома, пс – одинарная переходная спираль, 2пс – двойная переходная спираль, пф – переходные фибриллы.
стоящее время описано не более 10 истинно одножгутиковых видов.
Кинетосомы часто бывают связаны между собой фибриллярными мостиками и окружены электронно-плотным веществом, от которого берут начало их корешки. Базальные тела могут располагаться параллельно друг другу, особенно это характер но для многожгутиковых протистов и инфузорий, под острым, прямым или тупым углами, а также бывают развернуты под углом 180° (так называемое антипараллельное положение кинетосом). П о- казано, что у некоторых зеленых водорослей даже в течение онтогенеза клетки возможна переориентация кинетосом, скаж ем,
ñпараллельного положения на антипараллельное.
Корешковая система
Корешками обычно считаются фибриллярные и микротру-
бочковые структуры, котороые отходят непосредственно от кинетосом, или соединяются с ними короткими фибриллярными
162

4.2. ЦИТОСКЕЛЕТ
Рис. 4.23. Строение переходной зоны жгутика у воротничковых жгутиконосцев на продольном (А) и поперечном (Б) срезах. (Ориг.).
ак – аксонема, кн – кинетосома, пф – переходные фибриллы, цф – центральный филамент.
связками, или начинаются в слое аморфного материала, окру - жающенго базальное тело жгутика (рис. 4.14). Фибриллярные корешки могут быть поперечно-исчерченными и простыми. Про - стые, или неисчерченные, корешки встречаются сравнительн о редко. Среди поперечно-исчерченных корешков различают фи б- риллы системы I, которые имеют короткий период исчерченно с- ти (25–35 нм), состоят из структурного белка ассамблина, несократимы и обычно ассоциированы с микротрубочками; и фибриллы системы II с периодом исчерченности более 80 нм, которые образованы другим белком - центрином, - способны медленно сокращаться и не связаны с микротрубочками.
Микротрубочковые корешки образованы лентами или пуч- ками микротрубочек, а иногда и одиночными микротрубочками. Их строение и взаимное расположение весьма разнообраз - но (рис. 4.24–4.27).
Оба типа корешков выполняют прежде всего ту же функцию, которую выполняют корни у растения, закрепляя жгутик в те - ле клетки. Об этом свидетельствует тот факт, что жгутики без корешков не встречаются. Даже в тех исключительно редких случаях, когда они не обнаружены (Pelagomonas), следует предполагать, что они просто не выражены морфологически (напр и- мер, представлены многочисленными одиночными филамента -
163
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
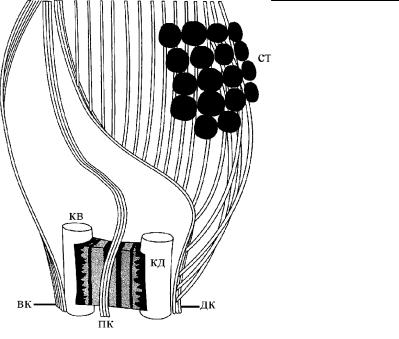
Рис. 4.24. Строение корешковой системы у эвгленовых. (По: Inouye, 1993.)
вк – вентральный корешок (направляется к рото-глоточным структурам клетки), дк – дорсальный корешок с отходящими о т него вторичными микротрубочками, которые фиксируют положение глобул глазка, или стигмы (ст), пк – промежуточный корешок (выстилает жгутиковый резервуар), кв – кинетосома вентрального жгутика связана фибриллярн ым мостиком с кинетосомой дорсального жгутика (кд).
ми, которые не заметны на ультратонких срезах). Об этом свидетельствуют многочисленные факты изоляции цитоскелета у самых разных протистов. При разрушении клетки детергентами ядро остается связанным с кинетосомами, независимо о т того, обнаружены ядерные корешки на ультратонких срезах или нет. Более того, при утрате кинетосом вместе с корешками жгутик может прикрепляться своим основанием к каким-либо другим структурам клетки. Удивительный случай описан при делении клетки зеленой водоросли Chlorogonium elongatum
164
4.2. ЦИТОСКЕЛЕТ
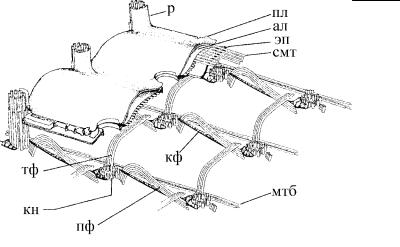
Рис. 4.25. Обобщенная схема строения соматического кортекса у инфузорий. (По: Margulis et al., 1993.) Передний конец клетки находится слева.
ал – альвеолы пелликулы, кн – кинетосома, кф – кинетодесмальный филамент (из микрофиламентов), мтб – лента базальных микротрубочек, пл – плазмалемма, пф – постцилиарная фибрилла (из микротрубочек), р – ресничка, смт – субпелликулярные микротрубочки, тф – трансверсальн ая фибрилла (из микротрубочек), эп – эпиплазма.
(Hoops, Witman, 1985). При подготовке к митозу кинетосомы этого протиста покидают свои жгутики и мигрируют вместе с корешками к полюсам ядра, где в дальнейшем участвуют в его де - лении как центриоли. Параксиальный конец аксонемы (на уровне переходной зоны) прикрепляется вновь образованными фибриллярными корешками к поверхности митохондрии, кото - рая выдвигается в апикальную зону клетки. Таким образом, оставшиеся без кинетосом жгутики продолжают активно работать, а клетка не меняет своего двигательного поведения и сохра няет те же фототактические реакции, что и перед делением.
Микротрубочковые корешки выполняют цитоскелетную функцию. Проходя под покровами или в глубь цитоплазмы клетки, они определяют и поддерживают ее форму. Это особен - но важно для организмов с тонкими покровами, не способными поддерживать, скажем, вытянутую форму клетки. Напри-
165
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Рис. 4.26. Основные типы организации жгутикового аппарата у зеленых водорослей. (По: Inouye, 1993.)
Левая колонка – вид сбоку, правая колонка – с переднего кон ца клетки. А – хлорофитовые, Б – ульвовые, В – требоуксиевые, Г – харовые. кн – кинетосомы, мсс – многослойная структура, у – узкий корешок, ф – фибриллярные связки между кинетосомами, фм – фибриллярный материал, ассоциированный с узкими микротрубочковыми корешками, ш – широкий корешок.
166

4.2. ЦИТОСКЕЛЕТ
Рис. 4.27. Многообразие корешковых систем жгутиков у охрофитов. (По: Inouye, 1993.)
Представлено в виде возможного филогенетического древа , построенного на основании сиквенсов генов РНК малой субъединицы рибосом. А – кинетосома переднего жгутика, Р – кинетосома заднего жгутика, номерами 1–4 обозначены гомологичные микротрубочковые корешки.
167

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
мер, клетка бодонид (кинетопластиды) покрыта плазмалеммо й, однако ее передний конец имеет глубокую инвагинацию в вид е жгутикового кармана и выступ в виде рострума. И та, и другая структура укреплены микротрубочковыми корешками.
Показано, что у гамет харовых водорослей они выполняют и морфогенетическую функцию. Так, во время созревания гаметы, которая исходно имеет форму шара, в ней вырастает широкий корешок из микротрубочек, идущий вдоль клетки. По мере роста корешка клетка вытягивается, а затем спирально закр у- чивается вокруг продольной оси, что также определяется из менением формы корешка при его дальнейшем развитии.
Наиболее развитая система корешков свойственна крупным жгутиконосцам и инфузориям, а также тем протистам, у котор ых длина двигательного жгутика значительно превышает длин у тела. Степень развития корешков зависит также от наличия ил и отсутствия аппарата питания (губа, цитостом, глотка и другие ).
Перед делением клетки происходит удвоение кинетосом, которые затем расходятся в дочерние особи. Детали этого про цесса пока еще слабо изучены. Показано, что для некоторых хризоф и- товых и зеленых водорослей с двумя жгутиками характерен п о- луконсервативный тип деления кинетиды (рис. 4.28). Сначала рядом с каждой кинетосомой формируется дочерняя. Затем об е пары кинетосом расходятся в дочерние клетки и после переориентации занимают свое окончательное положение. При этом к а- ждая дочерняя клетка наследует одну «старую» кинетосому и одну «новую», образовавшуюся перед митозом в результате удвоения. При этом кинетосома переднего жгутика материнс кой клетки становится кинетосомой заднего жгутика дочерней клетки, а кинетосома переднего формируется заново.
Консервативный тип деления кинетиды описан пока только для миксомицета Physarum polycephalum (Wright et al., 1980). В одной дочерней клетке кинетосома переднего жгутика ост а- ется прежней, а в другой клетке кинетосома заднего жгутик а становится кинетосомой переднего. Вновь образовавшиеся кинетосомы сначала формируют кинетосомы задних жгутиков, а в следующей генерации клеток становятся базальными тела ми передних жгутиков.
168

4.2. ЦИТОСКЕЛЕТ
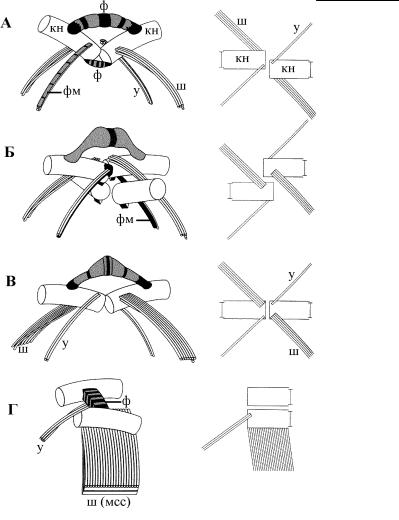
Рис. 4.28. Цикл развития жгутиков у зеленой водоросли Spermatozopsis. (По: Lechtreck et al., 1997.) А – интерфазное состояние, Б – удвоение кинетосом перед делением ядра, В – переориентация кинетосом во время деления клетки, Г – формирование дочерних кинетид на поздней стадии деления клетки. нст – новая стигма, ст – стигма, у – узкий микротрубочковый корешок, ш – широкий микротрубочковый корешок. 1, 2 – кинетосомы в интерфазной клетке, 21, 22 – дочерние кинетосомы, 12 – кинетосома 2 родительской клетки, которая превращается в кинетосому 1 дочерней клетки.
Поведение жгутиков может быть различно. У одних видов они теряются (втягиваются в клетку или отбрасываются) пер ед началом митоза, а после его завершения вырастают вновь. У других жгутики не исчезают, а наоборот, вновь образовавшиеся кинетосомы сразу дают новые жгутики, которые присутств у- ют на всех стадиях деления клетки.
Кинетосомы (или их корешки) многих жгутиконосцев могут участвовать и в делениии ядра в качестве ЦОМТов митотического веретена. Жгутики при этом обычно исчезают, а пары кинетосом мигрируют к полюсам делящегося ядра.
Гомология корешков
Большое значение для выяснения родственных отношений между протистами имеет установление гомологичных струк тур
169

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
(в частности, корешков). Первоначально гомология корешков была установлена у инфузорий. Их кинетида обычно характеризуется наличием трех корешков: одного фибриллярного (к и- нетодесмальный филамент) и двух микротрубочковых (транс - версальная фибрилла и постцилиодесма) (рис. 4.29). В пределах этого таксона была прослежена трансформация дериватов к и- нетосом и показаны пути их эволюции. В результате была произведена ревизия всей макросистемы инфузорий, опиравшей - ся ранее на признаки ротовой цилиатуры, и предложена нова я классификация. Интересно, что сходные результаты при построении системы инфузорий были независимо получены в США Д.Линном (D. Lynn) и в России Л.Н.Серавиным и З.П.Герасимовой примерно в одно и то же время (1976–1981 годы).
Âконце 70-х была продемонстрирована гомология микротрубочковых корешков у зеленых водорослей (рис. 4.26) и жгутиковых клеток высших растений (Stewart, Mattox, 1975, Moestrup, 1978, Melkonian, 1982, Sluiman, 1983), что привело,
âконечном счете, к перестройке всей системы зеленых водо - рослей и признанию монофилетичности всей группы: Chlorophyta, Charophyta и Plantae.
Позднее были получены доказательства гомологии дорсального и вентрального корешков у эвгленовых и кинетопластид. У тех и других они расположены латерально, начи- наются соответственно от кинетосомы переднего и заднего жгутиков и укрепляют стенки жгутикового кармана или резервуара (Brugerolle et al., 1979, Kivik, Walne, 1984, Frolov, Karpov, 1995).
Âначале 90-х была показана гомология микротрубочковых корешков у хризофитовых и близких к ним гетероконтных водорослей и зооспоровых грибов (Andersen, 1991). Для каждой кинетосомы характерно наличие пары микротрубочковых ко - решков, причем от кинетосомы опушенного жгутика обычно отходит ребристый корешок (рис. 4.27).
Âпоследнее время показана гомология корешков у церкомонад, протостелид и миксогастриевых (Karpov, 1997). В настоящее время усиленно ведутся поиски гомологичных коре ш- ков, объединяющих все группы протистов (Moestrup, 2000).
170