

Строение клетки протистов
.pdf4.2. ЦИТОСКЕЛЕТ
Рис. 4.29. Организация кинетиды и кинетома у инфузорий. (По: Lynn, Small, 1981.)
А – структура соматической кинетиды, Б – строение соматических кинет, В – организация кинетома (соматической и оральной цилиатуры). кд– кинетодесмальный филамент, кн – кинетосомы соматической цилиатуры, оц – оральная цилиату ра, пц – постцилиарная фибрилла, т – трансверсальная фибрилла .
Понятие кинетиды
Для общего определения жгутикового/ресничного аппарата часто используется понятие «кинетида». Определение этог о понятия для протистов не вполне четкое, т.к. первоначально было разработано только для инфузорий. Весь двигательный аппа - рат инфузорий обычно называется кинетом. Он состоит из ря - дов ресничек, которые определяются как кинеты. Кинетида ж е представляет собой «элементарную» структурно-функциона льную единицу кинеты или всего кинетома (рис. 4.29). Для жгутиконосцев, имеющих 1–2 жгутика, понятие кинетома и кинетиды совпадают. Наиболее консервативная часть кинетиды – кинетосомы, количество которых в кинетиде подавляющего большинства протистов равно двум. Двухкинетосомальное состояние кинетиды распространено наиболее широко, хотя есть
171

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
и истинно одножгутиковые формы (см. выше) и некоторые многожгутиковые (опалиниды), у которых кинетосомы непарные.
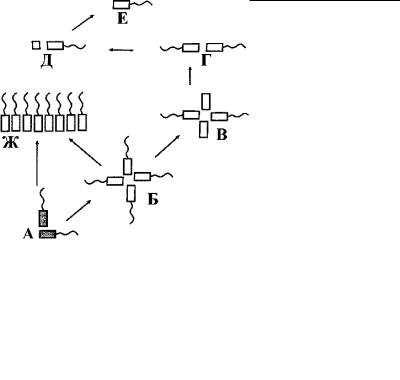
Для полимастигин (дипломонады, оксимонады, парабазалии) характерно объединение жгутиков с ядром в единый мор - фо-функциональный комплекс (рис. 4.30). Его принято называть кариомастигонтом. Обычно кариомастигонт содержит 4 жгутика и 4 кинетосомы, объединенные попарно. Поскольку пары кинетосом часто не связаны друг с другом морфологиче - ски, можно считать, что кариомастигонт содержит две кинет и- ды и одно ядро. Такая ситуация характерна для многих четырехжгутиковых протистов. По-видимому, такой кинетом сформировался в процессе эволюции путем удвоения двухки - нетосомальной кинетиды.
У протистов наблюдаются как процессы редукции двухкинетосомальной кинетиды, так и ее полимеризация. Их можно проследить в рамках того или иного монофилетичного таксо на. Так, было показано, что исходной для зеленых водорослей является двухкинетосомальная кинетида. В результате ее удвое-
Рис. 4.30. Схема строения переднего конца тела трихомонады
Ditrichomonas honigbergii. (По: Farmer, 1993.) аг – аппарат Гольджи, ак – аксостиль, п – пельта, пф – парабазальные фибриллы, р – кинетосома рекуррентного жгутика, ум – ундулирующая
мембрана, я – ядро. Пронумерованы кинетосомы, от которых отходят жгутики.
172
4.2. ЦИТОСКЕЛЕТ
ния появился наиболее распространенный среди них четыре х- жгутиковый тип кинетиды, подвергшийся в дальнейшем процессам редукции как жгутиков, так и кинетосом вплоть до ис - тинно одножгутиковой кинетиды Micromonas (рис. 4.31). Эта же двухкинетосомальная кинетида дала начало и кинетому г а- мет и зооспор эдогониевых водорослей, насчитывающему десятки жгутиков (Pickett-Heaps, 1971).
Рис. 4.31. Предполагаемые пути эволюции кинетиды у зеленых водорослей. (По: Карпов, 1993.)
А – исходная двухкинетосомальная кинетида (современные спермии харовых водорослей), Б – удвоение кинетиды (современные празинофитовые – Pyramimonas), В – редукция жгутиков (современные празинофитовые), Г – редукция кинетосом (современные Chlamydomonas), Д – полная редукция жгутика и частичная редукция одной из кинетосом (Pedinîmonas), Е – полная редукция одной из кинетосом (Micromonas), Ж – полимеризация жгутиков (современные зооспоры и спермии эдогониевых).
Полимеризация кинетиды у полимастигин (дипломонад, оксимонад и парабазалий) – генеральная линия эволюции. Исходным для них, по-видимому, должен считаться кариомастигонт с двумя парами кинетосом, которые тесно связаны с ядр ом (рис. 4.32). У дипломонад происходит удвоение кариомасти-
173
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
гонта, что приводит к появлению двухлучевой или даже била - теральной симметрии у особей, обладающих двумя ядрами и 8 жгутиками. Дальнейшее развитие этого пути полимеризации кариомастигонта находит отражение у оксимонад (рис. 4.32). В некоторых родах оксимонад число кариомастигонтов дост и- гает сотен на одну клетку.
Такой же тип полимеризации всего кариомастигонта характерен также для трихомонад из отряда Calonymphyda. Для большинства же их отмечена полимеризация только кинетиды, что имеет место у многожгутиковых гипермастигин с одним ядро м (рис. 4.32).
Недавно была обнаружена свободноживущая анаэробная трихомонада Ditrichomonas honigbergii (Farmer,1993), ó êîòî-
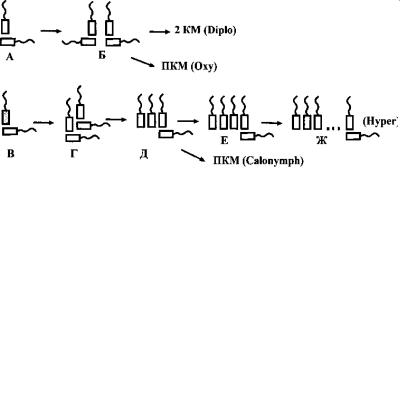
Рис. 4.32. Предполагаемые пути полимеризации кинетиды у полимастигин. (Ориг.).
А–Б – путь полимеризации кинетиды у дипломонад и оксимонад. А – исходная двухкинетосомальная кинетида (гипотети - ческая), Б – удвоение кинетиды (ретортамонады), что приводи т в дальнейшем к образованию устойчивой связи кинетиды с ядром и формированию двух кариомастигонтов (2 КМ) у дипломонад и поликариомастигонтов (ПКМ) у оксимонад.
В–Ж – путь полимеризации кинетиды у парабазалий. В – исходная двухкинетосомальная кинетида (гипотетическая ), Г – удвоение кинетиды (Ditrichomonas), Д – переориентация кинетосом, что приводит к образованию устойчивого кариом астигонта (трихомонады) и дальнейшей его полимеризации с формированием поликариомастигонта (ПКМ) у калонимфид, Е – увеличение числа передних жгутиков (трихомонады), Ж – полимеризация кинетиды (гипермастигины).
174

4.2. ЦИТОСКЕЛЕТ
рой 4 кинетосомы расположены попарно (рис. 4.30). Это попарное расположение кинетосом указывает, по-видимому, как
èу других полимастигин, на исходное двухкинетосомальное состояние кинетиды у предков всех полимастигин.
Полимеризация кинетиды от 2 исходных кинетосом отмечается также у протеромонад и ведет к многожгутиковости опалинат. Наиболее яркий пример такого типа полимеризации кинетид ы демонстрируют инфузории (рис. 4.29). Их кинетом формируется на основе полимеризации двух кинетосом (Lynn, 1991) и подвергается в дальнейшем глубокой морфологической и функционал ь- ной дифференцировке, часто ведущей к олигомеризации кине тома (Догель, 1951).
Обобщая, можно заключить, что полимеризация кинетиды у протистов чаще всего начинается с бикинетосомального со стояния, и первый этап этой полимеризации в подавляющем боль - шинстве случаев ведет к удвоению кинетиды. В дальнейшем, если устанавливается прочная связь с ядром и формируется кариомастигонт, полимеризации подвергается весь кариомастигонт, что приводит к появлению многоядерных и многожгутиковых протистов. Если же связь с ядром непрочная, полиме - ризация кинетиды идет независимо от ядра.
Принято считать, что поскольку бикинетосомальное состоя - ние наиболее широко распространено среди протистов, то он о
èявляется наиболее древним. Как мы видели, оно действительно наиболее консервативно, т.к. почти во всех группах протистов полимеризация начинается именно с 2 кинетосом. Кинето - сомы и центриоли имеют одинаковое строение, поэтому и происхождение кинетосом легко объяснить, если предполож ить, что центриолярный аппарат клетки был первичен. Тогда в эволюционном смысле кинетосомы – это центриоли, у которых выросли жгутики. Это наиболее общепринятая в настоящее вр е- мя точка зрения на происхождение жгутиков и ресничек.
Однако помимо этого широко представленного пути полимеризации кинетиды у протистов имеет место и другой, идущий , по-видимому, на основе одной кинетосомы (Карпов, 1993). Он характерен для некоторых хитридиевых грибов, протостели д (Myxomycetes) и шизопиренид (Heterolobosea). Их жгутико-
175

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
вые стадии имеют, как правило, непостоянное для вида число кинетосом, при этом каждая кинетосома обладает индивидуа льным набором корешков. Эти примеры свидетельствуют о возможном пути становления кинетиды протистов на основе однокинетосомального состояния.
Такое положение противоречит общепринятым представлениям о происхождении кинетиды из центриолей. Однако и его нельзя исключать из возможных путей становления кинетид ы у эукариот. В конце концов, появление центриолей тоже должно быть объяснено. Кроме того, эволюционно консервативные состояния (в данном случае бикинетосомальное) не обязате льно должны быть наиболее древними, а предковая форма (однокинетосомальное состояние) может встречаться весьма ред ко. Поэтому следует признать вполне вероятным и путь становл е- ния кинетиды эукариот на основе однокинетосомального со стояния кинетиды.
4.2.2. Функции жгутика/реснички
Основная функция жгутика – движение. В активной работе жгутика движущим началом являются периферические микротрубочки и их динеиновые ручки, обладающие АТФ-азной активностью. На изолированных дублетах микротрубочек ак - сонемы показано, что при добавлении АТФ происходит скольжение дублетов относительно друг друга. Находясь в состав е жгутика, дублеты связаны друг с другом и с центральными микротрубочками, формируя длинный цилиндр. Поэтому скольжение дублетов относительно друг друга, вызванное работой динеиновых ручек, приводит к изгибанию аксонемы. Централь - ные микротрубочки выполняют при этом опорную функцию. Исследования мутантов Chlamydomonas reinhardtii показали, что внутренние динеиновые ручки вызывают изгибание жгутика и определяют размер и форму волны биения. Наружные ручки усиливают этот эффект и увеличивают частоту биения . Недавно было установлено, что центральные микротрубочки выполняют не только опорную функцию, но вместе с радиаль-
176

4.2. ЦИТОСКЕЛЕТ
ными спицами взаимодействуют с внутренними ручками, контролируя форму биения жгутика (Porter, Sale, 2000).
Формы движения жгутика различны, но обычно это волнообразное изгибание с частотой до 50 колебаний в секунду. У большинства протистов биение жгутика происходит в одной плоскости. У инфузорий и многожгутиковых протистов движе - ние ундулиподий организовано по типу метахрональных вол н. При этом биение отдельной реснички состоит из двух этапов : рабочий удар и возвратное движение (рис. 4.33). Во время рабочего удара прямая ресничка совершает гребное движение спереди назад, а при возвратном движении она мягко волнообра з- но изгибается, возвращаясь назад уже по другой траектории (рис. 4.33), максимально приближенной к поверхности тела, и сводя к минимуму тормозное влияние. Таким образом, рабо- чий удар ресничек направляет движение клетки в противопо - ложную сторону.
Рис. 4.33. Кинетика биения ресничек и жгутиков. (По: Sleigh, 1991.)
При рабочем ударе (А) ресничка захватывает бульшую массу воды (ограничено пунктирной линией), чем при возвратном (Б) движении. В – форма биения жгутиков у плывущей хламидомонады. Стрелками показано рабочее движение жгутиков.
177

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Следует подчеркнуть, что с точки зрения физики передвижение в жидкости малых объектов, какими являются жгутиконосцы и инфузории, существенно отличается от движения крупных организмов (например, рыб). Вообще к движущемуся в воде или воздухе телу приложимы следующие силы: силы инерции движущегося тела и силы вязкости, связанные со скольжением слоев жидкости относительно друг друга. Отно - шение сил инерции к силам вязкости выражается числом Рейнольдса (R), которое прямо пропорционально размерам движущегося тела. Например, для крупной рыбы число R может составлять 1000, тогда как для жгутиконосца оно будет менее 0,01. Таким образом, если для рыбы силы вязкости пренебрежительно малы по сравнению с силами инерции, то для жгутиконосца, наоборот, силы инерции в сто раз меньше сил вязкости, и уже силами инерции можно пренебречь. Другими словами , как только жгутики или реснички перестают работать, клетк а сразу же останавливается.
По сравнению с размерами тела клетки, прилегающий к ее поверхности ламинарный слой жидкости имеет значительну ю толщину. Поэтому во время рабочего удара расправленная ре с- ничка захватывает и перемещает сравнительно большой объ ем жидкости (рис. 4.33). Во время возвратного движения реснич- ка пригибается к поверхности клетки и проходит вблизи нег о, поэтому объем захватываемой и перемещаемой жидкости зна - чительно меньше, чем при рабочем ударе (рис. 4.33). Это и обеспечивает поступательное движение клетки. Метахрональны е волны, пробегающие по поверхности тела многожгутиковых и ресничных протистов, могут быть организованы различным о б- разом. В зависимости от направления рабочего удара по отн о- шению к направлению распространения метахрональной вол - ны различают симплектическую (рабочий удар направлен по ходу метахрональной волны), антиплектическую (рабочий уд ар направлен против волны) и диаплектическую (рабочий удар н а- правлен под углом к волне) формы метахронии (рис. 4.34).
Диаплектическая форма метахрональной волны, в свою оче- редь, делится на 2 вида (рис. 4.34): леоплектическую (рабочий удар реснички направлен влево, если смотреть по ходу расп ро-
178

4.2. ЦИТОСКЕЛЕТ
Рис. 4.34. Основные формы биения ресничек. (По: Sleigh, 1989.)
Показана связь между направлением рабочего удара реснич ки (тонкая стрелка) и направлением метахрональной волны (толстая стрелка). На рис. В и Г (диаплектическая форма) рабочий удар реснички направлен на читателя. А – симплектическая форма, Б – антиплектическая, В – леоплектическая, Г – дексиоплектическая.
странения волны) и дексиоплектическую (рабочий удар реснички направлен вправо, если смотреть по ходу распростран е- ния волны). Большинство ресничных полей у инфузорий демонстрируют диаплектическую форму метахронии.
Жгутики и реснички часто используются также и для питания. При этом форма биения жгутика может меняться по сравнению с двигательным, а может оставаться неизменным. Формы биения жгутиков, вызываемых ими токов жидкости и направления движения клетки у различных протистов показ а- ны на рисунке 4.35. Помимо движения и питания протисты используют жгутики для того, чтобы очищать поверхность тела от налипающих на него мелких посторонних частиц.
Многие хризомонады имеют редуцированный рулевой жгутик. Обычно в его основании расположено вздутие, представ - ляющее часть фоторецепторного аппарата, т.е. жгутики могут выполнять и рецепторную функцию.
179

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Рис. 4.35. Направление движения воды, вызываемое биением ресничек и жгутиков. (По: Sleigh, 1989.)
А – хризомонада Ochromonas, Б – воротничковый жгутиконосец Codonosiga, В – инфузория Euplotes, Г – инфузория Stentor.
Среди жгутиконосцев имеются виды, проводящие большую часть жизненного цикла в прикрепленном состоянии. В этот период жгутик теряет обычную для него функцию движения и превращается в органеллу прикрепления: стебелек или ножку. Это характерно как для свободноживущих (рис. 2.32), так и для паразитических (рис. 4.36) форм.
Рис. 4.36. Схема прикрепления трипаносомы к щеточной каемке кишечного эпителия при помощи видоизмененного жгутика. (По: Фролов, Скарлато, 1997.)
ж – жгутик, жр – жгутиковое расширение, мв – микровилли клетки кишечного эпителия, тк – тело клетки трипаносомы.
180