

1 0 0 Г Л А В А 1 1 . Р О Д И Т Е Л Ь С К И Е Э Ф Ф Е К Т Ы
Сходное явление было открыто у цветковых растений. Второе отдельное оплодотворение приводит к образованию эндосперма, экстраэмбрионной структуры, которая питает семя подобно плаценте млекопитающих. Эндосперм часто содержит две копии материнского генома и одну отцовского, что приводит к триплоидному геному (2 ♀: 1 ♂). Избыток материнских генов (соотношение больше чем 2 ♀: 1 ♂) приводит к угнетению развития эндосперма, тогда как избыток отцовских генов как правилокоррелируетсизбыточнымростом(Haig, Westoby, 1991).
Теориигеномногоимпринтинга
Предотвращение партеногенеза. Одним из следствий геномного импринтинга является предотвращение партеногенеза. Тогда можно ожидать, что он не должен встречаться в группах размножающихсяпартеногенезом.
Теорияродительскогоконфликта(The parental conflict hypothesis). ГипотезаХейга(Haig, Westoby, 1989) объясняет геномный импринтинг конфликтом интересов между матерью и отцом за количество ресурсов, передаваемых от матери к плоду. Для растений это выражается в том, что эндосперм (ткань отвечающая за накопление ресурсов) является возможным местом проявления геномного импринтинга. У животных плацента также выступает как основное “поле битвы”, при этом отцовские гены должны увеличивать экстракцию питательных веществ во время беременности, тогда как материнскийгеномдолженеёограничивать.
"Отцовский" эффект
“—Неужелинеузнаете? Амеждутеммногиенаходят, чтояпоразительнопохожнасвоегоотца.
—Ятожепохожнасвоегоотца,—нетерпеливосказалпредседатель.
—Вамчего, товарищ?” “Золотойтеленок” (И. Ильф, Е. Петров, 1993)
Поскольку предлагаемая концепция рассматривает половой диморфизм как следствие гетерохронной эволюции признаков у мужского и женского пола, как эволюционную “дистанцию” между полами, то естественно ожидать максимальных проявлений полового диморфизма по эволюционирующим (новым) признакам. У человека к таковым, видимо, можно отнести в первую очередь всё, что связано с корой головного мозга, а у сельскохозяйственных животных и растений—все хозяйственно-ценные признаки. Поэтомуонирассмотреныболеедетально.
Как уже говорилось в гл. 3, в связи с полом можно выделить три группы признаков: признаки по которым половой диморфизм в норме отсутствует, признаки, характерные только для одного пола, и признаки, выраженыевразнойстепениуобоихполов.
О генотипическом половом диморфизме по признакам, присущим только одному полу можно судить по реципрокным эффектам. Если по “старым” (стабильным) признакам генетический вклад отца в потомка в среднем несколько меньше вклада матери из-за материнского эффекта, обусловленного цитоплазматической наследственностью, гомогаметной конституцией и утробным развитием у млекопитающих, то по “новым” признакам, согласно эволюционной теории пола, вклад отца должен бытьбольше. Это может привести к компенсации материнского эффекта по таким признакам и дажек появлению противоположного ему “отцовского” эффекта. Иными словами, при передаче генетической информации по “новым” признакам должно существовать некоторое доминирование отцовскихпризнаковнадматеринскими.
При дивергентной эволюции всегда бывает стадия, когда мужской пол, как авангард, уже бимодален, аженский—арьергард, ещемономодален, тогдаобагибрида, двухразныхсамцовсобщейсамкой, при аддитивном наследовании, (♀+♂)/2, попадают в вид отца. А приконвергентной эволюции двух видов,

Г Л А В А 1 1 . Р О Д И Т Е Л Ь С К И Е Э Ф Ф Е К Т Ы 1 0 1
самцы по новому признаку уже одинаковы, а самки ещё разные и оба гибрида попадают в вид матери. Поскольку эволюция в многомерных нишах (тропики) и вся селекция дивергентна, отцовский эффект наблюдается намного чаще, материнского. А конвергентная эволюция бывает в одномерных нишах, гдедоминируетодинфакторсреды: мороззаполярнымкругом, илижаравпустыне.
ФИЛОГЕНЕТИЧЕСКОЕ ПРАВИЛОРЕЦИПРОКНЫХЭФФЕКТОВ
УРЕЦИПРОКНЫХГИБРИДОВПОДИВЕРГИРУЮЩИМПРИЗНАКАМ РОДИТЕЛЕЙДОЛЖНАДОМИНИРОВАТЬОТЦОВСКАЯФОРМА(ПОРОДА), АПОКОНВЕРГИРУЮЩИМ —МАТЕРИНСКАЯ.
Реципрокный “отцовский” эффект позволяет отличить эволюционирующий признак от стабильного. Но он ничего не говорит о направлении эволюции признака, о котором можно судить по
генотипическому половому диморфизму и гетерозису. Рассматривая явление гетерозиса как суммациюновыхэволюционныхдостижений, приобретенных дивергентно, можнопредположить, что вклад отца в гетерозис также должен превышать вклад матери. В свете новых представлений становится понятным почему при гетерозисе, как правило, усиливаются признаки полезные для человека, а не для самих претерпевших гетерозис видов, независимо от того, у кого наблюдается гетерозис: у кукурузы, томатов или кур. Самим этим видам гетерозис мало что дает, а может быть и вреден. Однако, поскольку селекцию можно рассматривать как навязанную человеком культурным видам искусственную эволюцию, то и вектор этой эволюции и вектор гетерозиса совпадают с утилитарными интересамичеловека, анесэволюционнымиинтересамиэтихвидов.
Далее, рассматривая эволюционное преобразование признака в филогенезе как некоторое абстрактное “движение”, можноговоритьо“расстоянии” междумужским иженскимполомпоэтому признаку.
Если исходная форма в филогенезе дивергировала на две отличающиеся по данному признаку формы
(породы, линии, расы) А и В, то согласно филогенетическому правилу полового диморфизма, можно ожидать, что в каждой из появившихся форм (А и В) мужской пол должен опережать женский пол по изменению этого признака. Значит, можно говорить о “расстоянии” между породами по этому признаку (насколько они удалились друг от друга) и о “расстоянии” между мужским и женским полом (Рис. 11.1). Следовательно, при гибридизации, в принципе, можно выделить “вклад” породы и “вклад” пола. О “расстоянии” между породами или о вкладе пород можно судить по явлению гетерозиса, а о “расстоянии” или вкладе полов—по половому диморфизму. А по направлению реципрокного эффекта (материнский или отцовский эффект) и его величине можно судить о дивергентном или конвергентном характере эволюции признака. Таким образом, появляется возможностьполнее объяснить реципрокные эффекты, которые естьнечто иное, как векторная сумма материнского иотцовскогоэффектов.
Рис. 11.1 |
|
|
|← |
Вклад породы |
|
|
Направление |
Оценкареципрокныхэффектов |
|
|
|
|
|
|
эволюции |
|
♀A |
♂A |
|
♀B |
♂B |
признака |
|
|
|
|
|||||
|
|
▼ |
▼ |
|
▼ |
▼ |
|
|
|
|← |
→| |
|
|← |
→| |
|
|
|
|
|
Вклад пола |
|
|
|
|
|
|
|
Скрещивания |
|
|
|
|
♂A x ♀A |
♂A x ♀B |
♂B x ♀A |
♂B x ♀B |
|||
|
|
|
|
Значение признака |
|
|
|
|
|
A |
|
a |
b |
|
B |
1 0 2 Г Л А В А 1 1 . Р О Д И Т Е Л Ь С К И Е Э Ф Ф Е К Т Ы
Дляизмеренияреципрокных эффектов( r ) можно использоватьвыражение:
r = (b - a) / (B - A) |
[ 5 ] |
где A и B —значения признака для исходных скрещиваемых форм; a —то же самое для гибрида ♂A x ♀B; b —для реципрокного гибрида ♂B x ♀A. Тогда положительное значение r (r > 0) будет означать “отцовский” эффект, отрицательное (r < 0)—материнский, а абсолютная величина r (│r│) даст относительную оценку этих эффектов в единицах, равных разности значения признака для исходныхформ(B - A).
Вектор “отцовского” эффекта по тому или иному признаку, так же как и вектор полового диморфизма, может служить “компасом”, показывающим направление эволюции этого признака. Но в отличие от полового диморфизма, который применим только для количественных признаков, присущих обоим полам по которым имеется половой диморфизм, “отцовский” эффект может показывать направление эволюции также и тех признаков, которые присущи только одному полу, в том числе и направление эволюции первичных половых признаков. Иными словами, реципрокный эффектдаетпринципиальнуювозможностьвыявитькакбыгенотипическийполовойдиморфизм.
У сельскохозяйственных животных и растений “эволюционно новыми”, очевидно являются все хозяйственно-ценные признаки. В Табл. 11.1 (Приложение В) приведены результаты разных авторов по наследованию признаков реципрокными гибридами кур, свиней и крупного рогатого скота. Робертс и Кард (Roberts, Card, 1933) изучали наследование инстинкта насиживания у реципрокных гибридов кур породы леггорн и бойцовых корнишей. У леггорнов в связи с промышленным (инкубаторным) разведением этот инстинкт практически утрачен. Среди бойцовых корнишей почти все куры клохчут. Признак клохтанья имеет существенное хозяйственное значение, так как куры, обладающие этим признаком, отличаются, как правило, меньшей яичной продуктивностью. Частота и степень проявления клохтанья была выше тогда, когда отцовской породойвыступалибойцовыекорниши.
Аналогичные результаты получили Морли и Смит (Morley, Smith, 1954), исследовавшие тот же признак у реципрокных гибридов пород леггорн и австралорп (у кур породы австралорп инстинкт насиживания присутствует), а также Саеки с соавт. (Saeki е. а., 1956) на реципрокных гибридах местной породы нагойя (клохчут почти все куры) с леггорнами. Как видим, при наследовании инстинкта насиживания у кур наблюдается четкий отцовский эффект (r = 0.45; 0.38 и 0.50 соответственно). По другому признаку—половой скороспелости кур (возраст снесения первого яйца)—исследовались межлинейные гибриды породы род-айленд, имеющей скороспелые и позднеспелые линии (Warren, 1934). По скороспелости наблюдается отцовский эффект r = 0.59. Сходные результаты были получены на межпородных гибридах австралийскими и японскими учеными (Morley, Smith, 1954; Saeki e. a., 1956) при скрещивании местных пород австралорп и нагойя (позднеспелыепороды) сболеескороспелойпородойлеггорн.
При наследовании еще одного хозяйственно-ценного признака—яйценоскости, по данным Уоррена (Warren, 1942) из пяти серий скрещиваний в четырех проявляется четкий “отцовский” эффект. Аналогичные исследования Нокса при скрещиваниях инбредных чистопородных леггорнов и родайлендов с 1946 по 1956 г., также подтверждают существование “отцовского” эффекта (цит. по Дубинин, Глембоцкий, 1967) (r = 1.14).
На Братцевской птицефабрике проводились опыты по определению яйценоскости, живого веса и веса яиц для пород московская и леггорн и их реципрокных гибридов. По яйценоскости и живому весу наблюдается совершенно отчетливый “отцовский” эффект, а по весу яиц—материнский, то есть, как и следовало ожидать, яйценоскость выше у дочерей отца-леггорна, а живой вес, наоборот,—у дочерей отцамосковскойпороды(Добрынина, 1958).
Г Л А В А 1 1 . Р О Д И Т Е Л Ь С К И Е Э Ф Ф Е К Т Ы 1 0 3
Большее влияние отца на яйценоскость дочерей объясняли тем, что у птиц гетерогаметным полом является самка, а гомогаметным—самец. Поскольку свою единственную Х-хромосому курица получает от отца, и если яйценоскость определяется ею, то тогда все понятно (Дубинин, Глембоцкий, 1967). Придерживаясь такой трактовки, следует ожидать, что у млекопитающих, у которых гетерогаметны самцы, все должно быть наоборот; независимо от того, наследуется “старый” или “новый” признак, должно наблюдаться большее влияние матери, чем отца, в то время как по эволюционной теории пола независимо от гаметности во всех случаях по эволюционирующим признакамдолженнаблюдаться“отцовский” эффект.
М. Асланян (1962) изучал наследование числа позвонков и некоторых характеристик пищеварительной системы у двух пород свиней и их реципрокных гибридов—шведский ландрас и крупная белая. Шведский ландрас—порода мясо-беконного направления. Она отличается от крупной белой—породы универсального мясо-сального направления более длинным туловищем и лучшим использованием корма, так как более полвека селекция велась на длинное туловище и более эффективную оплату корма. В 1909 г. количество кормовых единиц на 1 кг привеса у свиней породы ландрас составляло 4.5 кг, а в 1951 г.—3.7 кг. За то же время туловище у них удлинилось в среднем на 14 см. Этоудлинение происходило как за счет увеличения числа предкрестцовых позвонков (которых в среднем у ландрасов на 1.2 позвонка больше, чем у крупной белой породы), так и за счет удлинения самих позвонков. Кишечник лучше развит у ландрасов, главным образом за счет тонких кишок
(Асланян, 1962).
Позже изучение наследования количества позвонков у поросят (по рентгеновским снимкам) было проведено Б. В. Александровым (1966) на большем материале. Видно, что по наследованию числа позвонков у реципрокных гибридов свиней также наблюдается четкий “отцовский” эффект. Наследование различных характеристик пищеварительной системы выявляет “отцовский” эффект только по средней длине тонкого кишечника и пищевода на фоне материнского эффекта по другим признакам (среднему весу эмбрионов, пищеварительной системы и различных ее частей, длине толстого кишечника). “Отцовский” эффект наблюдается также и по динамике роста реципрокных гибридов (r = 1.8). Таким образом, “отцовский” эффект обнаруживается именно по тем признакам, по которым шла селекция ландрасов: числу позвонков (отбор на длинное туловище), длине тонкого кишечника (отбор на лучшую оплату корма) и динамике роста (отбор на скороспелость). Следует отметить, чтоповесу новорожденных поросятнаблюдаетсяматеринский эффект.
У трех пород крупного рогатого скота и их реципрокных гибридов по удою молока и продукции молочного жира также наблюдается предсказанный теорией “отцовский” эффект, хотя и не такой четкий, каквслучаях, описанных выше(Fohrman e. a., 1954).
Обратим внимание на наличие материнского эффекта по размеру яиц у кур и по проценту жиpa в молоке у коров. Оба признака являются хозяйственно-ценными, очевидно отбирались человеком в процессе доместикации и окультуривания и, казалось бы должны показать “отцовский” эффект. Если мы сравним размеры и число яиц у культурных пород кур и их диких предков, которые в природных условиях размножались сезонно и поэтому производили сравнительно малое количество яиц, но видимо, почти такого же размера, то увидим, что в результате окультуривания сильно увеличилось в первую очередь число яиц. Размерыяицизменились, видимо, мало.
Также, если сопоставим аналогичным образом процент жира в молоке и количество молока у культурных пород коров и их диких предков, которые производилималомолока, носвысокимсодержанием жира, тоувидим, что в результате селекции возросло количество молока, а процент жира даже снизился.
Современные куры несут по
126–200 яиц в год (рекорд—1515
яиц за 8 лет).
Продуктивность диких предков кур—5–14 яиц в год.
И. Акимушкин Мир животных. М., Молодая Гвардия, 1973, c. 207.
1 0 4 Г Л А В А 1 1 . Р О Д И Т Е Л Ь С К И Е Э Ф Ф Е К Т Ы
Наличие “отцовского” эффекта у млекопитающих позволяет считать существующую трактовку этого явления у кур (как следствие гетерогаметной конституции самок) по крайнеймеренедостаточной. Эта трактовка может объяснить хромосомный механизм явления у птиц, но для млекопитающих уже неприменима. Явление гораздо шире, оно связано не только с феноменом гетерогаметности, а и с феноменом пола и с эволюционными преобразованиями популяций. У млекопитающих оно реализуется, повсейвидимости, другими, отличнымиотгетерогаметности механизмами.
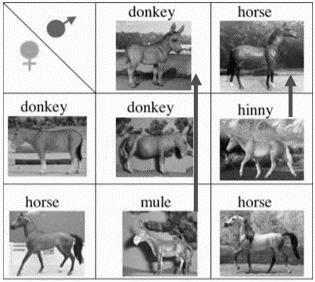
“Отцовский” эффект позволяет объяснить эпохальную загадку “асимметрии” родителей при реципрокных скрещиваниях, которую не может объяснить классическая генетика. Реципрокные гибриды лошади с ослом, появились ~8-10 тыс. лет назад, но до сих пор не понятно, почему и мул и лошакпохожинасвоихотцов(Рис. 11.2).
Рис. 11.2 Мулилошакпохожинасвоихотцов
Лошадь и осёл возникли дивергентно от общего предка тарпана (или лошади Пржевальского). Более далекие предки были величиной с кошку. После дивергенции лошадь стала длиннее, выше, быстрее, хвост из длинных волос. У осла удлинились уши, укрупнилась голова, сузились копыта, утончился хвостскистьюизволос. Геныэтихпризнаков, какновые, должныбыть вначалетолькоу самцов.
Таким образом приведенный материал позволяет считать, что вклад отца в гетерозис больше вклада матери и закономерность доминирования отцовских форм по эволюционирующим или селекционным у сельскохозяйственных животных признакам действительно имеет место. Генетическая информация по вторичным половым признакам (видимо, также и по первичным) присутствует и у мужского и у женского пола. “Отцовский” эффект наряду с генотипическим половым диморфизмом может служить свидетельством эволюции признака. Он позволяет качественно предсказывать результаты гибридизациииосуществлятьправильныйподборродительских парприскрещиваниях.
Геномный импринтинг является частным случаем более общего явления асимметрии родительских геномов, которая может проявляться как в форме присутствия – отсутствия генов, так и их экспрессии
– репрессии.