

Г Л А В А 7 . Д В А П О Л А — Д В А П О Т О К А И Н Ф О Р М А Ц И И |
7 5 |
Повышенный уровень мутаций у мужских особей
Поскольку число клеточных делений в сперматогенезе намного больше числа делений в овогенезе и ошибки при репликации и репарации ДНК являются главным источником мутаций, был сделан вывод, что все это может привести к большей частоте мутаций в половых хромосомах по сравнению с аутосомами и было предположено, что самцы служат генератором мутаций, по крайней мере, в эволюции млекопитающих. Тот факт, что уровень, как спонтанного так и индуцированного мутагенеза, у гетерогаметных, и у гомогаметных самцов выше чем у самок для дрозофилы, шелкопряда, млекопитающих, в том числе и человека был установлен давно и неоднократно
(Анисимов, Соловьев, 1999; Kerkis, 1975).
Сравнительным исследованием замещений нуклеотидной последовательности А-, Х- и Y-генов человека и мыши (или крысы) было показано, что самцы являются главным источником мутаций для молекулярной эволюции. Кроме того, было установлено, что соотношение скоростей эволюции генов— Y : А: Х= 2.2 : 1 : 0.6 (хорошосогласуетсястеоретическим ожиданием— 2 : 1 : 2/3) (Miyata e. a., 1987). Более высокий уровень точковых мутаций отмечен также у самцов мышей по сравнению с самками(Searle, 1972).
Наследование родительских признаков
Бадр и Спикет (Badr, Spickett, 1965) обнаружили различия между самцами и самками мышей в относительной роли аддитивных и неаддитивных наследственных факторов в детерминации веса надпочечников. Доля аддитивного генетического разнообразия в фенотипической изменчивости веса надпочечниковсоставляла 40% длясамцови60% длясамок.
По данным Л. Шюлера и др. (1976) у самок мышей гибридов значения относительного веса надпочечников, тимуса, половых желез и гипофиза практически совпадали с полусуммой значений этого признака у родителей. У самцов гибридов наблюдалось отклонение среднего в направлении меньшего значения для надпочечников, тимуса и половых желез, и большего—для гипофиза. У самок мышей гибридов было отмечено также более аддитивное наследование генов, ответственных за двигательнуюактивность(Бородин идр. 1976).
Более широкая норма реакции женского пола
Разная фенотипическая дисперсия полов может быть связана с их разной нормой реакции. Норма реакции, или модификационная изменчивость, особей женского пола должна быть больше соответствующей нормы реакции особей мужского пола. Иными словами, связь фенотипа с генотипом у мужского пола должна быть более “жесткая”, чем у женского. Поэтому влияние среды в онтогенезеприреализациигенотипавфенотипбольшеу женского пола(Рис. 7.2, Рис. 7.5).
В “пространстве возможностей” существует некая зона “онтогенетического дискомфорта”, находясь в которой особи не гибнут, но испытывают трудности и неудобства, страдают от неприспособленности (мерзнут, голодают и пр.). Широкая норма реакции позволяет особям женского пола покинуть эту зону и выжить, в то время как мужские особи остаются в ней и гибнут. У женских особей за счет широкой нормы реакции онтогенетическое “пространство способностей” больше, чем у мужских. Поэтому половую дифференциацию можно трактовать еще как специализацию по взаимно дополнительным качествам совершенства (женский пол) и новаторства, прогрессивности (мужской) во взаимоотношениях со средой, так как адаптация женских особей к существующим условиям среды совершеннее, чем у мужских особей. А у последних наличие разнообразных отклонений от нормы (моды) популяции (несовершенств) обеспечивает им большую прогрессивность в приспособлении к изменившимсяусловиямсредывбудущем.
7 6 Г Л А В А 7 . Д В А П О Л А — Д В А П О Т О К А И Н Ф О Р М А Ц И И
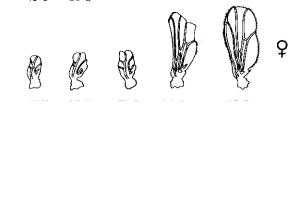
Рис. 7.5
Изменениеразмераиформыкрылау самцовисамокдрозофилывзависимости оттемпературысредысодержания
(Шмальгаузен, 1969). Вонтогенезеиз-заболее широкойнормыреакциивлияниесреды напроявлениепризнакаусамокбольше, чему
самцов, авлияниегенотипа, наоборот, меньше.
Узкая норма реакции обеспечивает широкую фенотипическую дисперсию мужского пола. Она дает возможностьподвергнутьэлиминациии дискриминации восновном мужскойпол. Приэтом, бóльшее сечение канала связи мужского пола с потомством позволяет максимально тиражировать отобранные, самыеподходящие, генотипы.
И наоборот, широкая норма реакции женского пола сужает его |
“ ...если объединить всех |
фенотипическую дисперсию и позволяет сохранить его |
самцов популяции в |
численность и существующее распределение генотипов. На |
мужскую команду, а всех |
первый взгляд кажется, что чем шире норма реакции, тем шире |
самок—в женскую и |
должна быть и фенотипическая дисперсия. Однако это не так. На |
устроить “личные и |
самом деле все наоборот: впопуляциифенотипическая дисперсия |
командные соревнования”, |
мужского пола, состоящего из особей с узкой нормой реакции, |
то чемпионами в “личном |
будет шире, так как она сохранит генотипическую дисперсию, а |
первенстве” по всем |
фенотипическая дисперсия женского пола, состоящего из особей |
программам были бы |
с широкой нормой реакции, будет уже, так как широкая норма |
самцы, а в “командном |
реакции даст возможность в условиях стабилизирующего отбора, |
зачёте” победителями |
крайним генотипам “уйти” из зон “дискомфорта”, “подтянуться” |
оказывались бы самки” |
к фенотипической норме (моде) популяции и таким образом |
|
сгладит генотипическую дисперсию (Рис. 7.2). Именно высокая |
Геодакян, 1966. |
онтогенетическая пластичность женского пола обеспечивает ему высокую стабильность в филогенезе. Следовательно, в
филогенетическом плане более изменчив, пластичен мужской пол, а в онтогенетическом, наоборот, более пластичен и изменчив женский. Такое, на первый взгляд парадоксальное, распределение ролей в филогенезе и онтогенезе на самом деле последовательно и непротиворечиво: оно реализует единую идею специализации полов по консервативной и оперативной задачам эволюции. Несколько утрируя, можно сказать, что информационные взаимоотношения популяции со средой строятся на элиминации мужских особейи“воспитании” (онтогенетическомсдвиге) женских.
Гипотеза более широкой нормы реакции женского пола позволяет делать довольно четкие предсказания. Например, у особей мужского пола доля “наследственной компоненты” должна быть больше, а “средовой”—меньше, чем у особей женского пола. Если сравнить внутрипарную и межпарную дисперсию у однояйцевых (монозиготных) близнецов мужского и женского пола, то внутрипарная должна быть больше у женских пар, а межпарная—у мужских, то есть степень конкордантности у однояйцевых близнецов мужского пола должна быть больше, чем у таких же близнецов женского пола. Далее, фенотипическая дисперсия в чистой линии может быть относительноширеуженского пола, авполиморфной(дикой) популяциионаширеумужского.
Ванденберг и др. (Vandenberg et al., 1962) исследовали внутрипарные различия у 23 пар мужских однояйцевых близнецови21 пары женских по различным измерениям: антропологическим, сердечно-
Г Л А В А 7 . Д В А П О Л А — Д В А П О Т О К А И Н Ф О Р М А Ц И И |
7 7 |
сосудистым, гематологическим, биохимическим, психологическим и др. Возраст исследованных пар от 12 до 18 лет. По 185 переменным внутрипарный разброс оказался статистически значимо больше у женских пар, по 41—у мужских, а по 353 переменным не было обнаружено значимой разницы. Аналогичныерезультатыбылиполученыидругимиавторами(Никитюк, 1977; Chovanova, 1980).
Исследование степени разнообразия по 9 параметрам (социальные, поведенческие и умственные способности в возрасте 2, 3 и 4 года) у 2000 пар однояйцевых близнецов показало большее разнообразие в парах девочек Интересно, что в полном соответствии с представлениями теории у дизиготныхблизнецовнаблюдаласьобратнаякорреляцияпотемжепризнакам. (Loat et al., 2004).
Шюлер Л. и др. (1976) обнаружили различия между самцами и самками мышей в детерминации веса эндокринных органов. Доля генотипической компоненты в фенотипической изменчивости у самцов по всем изученным признакам была больше, чем у самок, что также подтверждает гипотезу нормы реакции.
Более широкая фенотипическая дисперсия мужского пола выявляется в различных тестах по определению коэффициента интеллектуальности (IQ). У женщин показатели IQ группируются вокруг некоторых средних величин, а у мужчин—наблюдается значительный разброс по этому показателю
(Hurt, 1978).
Психологи |
|
из |
Эдинбургского |
университета |
проверили |
“Мужчины либо очень |
|||
интеллектуальный уровень у более чем 2500 братьев и сестер, |
умны, либо уж очень |
||||||||
подвергнув их тестам по информатике, математике и английскому, а |
глупы”: |
||||||||
также оценив их способность к выполнению механических |
...среди умных двух |
||||||||
операций. |
Мужчины набрали |
максимальное |
и |
минимальное |
процентов населения |
||||
количество очков по всем тестам, они также лучше справились с |
|||||||||
мужчин в два раза |
|||||||||
заданиями |
по |
информатике и |
арифметике. Женщины получили |
||||||
больше, чем женщин. |
|||||||||
средние оценки и выглядели лучше в языковых тестах. В два раза |
|||||||||
|
|||||||||
больше мужчин чем женщин оказалось на периферии |
... среди наименее |
||||||||
распределения—как среди двух процентов умных, так и среди двух |
интеллектуальных |
||||||||
процентов |
наименее |
интеллектуальных испытуемых |
(Deary, et al., |
двух процентов |
|||||
2007). |
|
|
|
|
|
|
|
населения мужчин в |
|
Американские |
исследователи |
|
проверяли |
математические |
два раза больше, чем |
||||
|
женщин. |
||||||||
способности (SAT тест) учеников 7-го класса на крайней правой |
Deary, e.a., 2007. |
||||||||
границе распределения (верхний 0.01 % популяции). В начале 80-х годов на каждую девочку приходилось 13 мальчиков. К 1991 г.
разница уменьшилась до 4 : 1 и оставалась на этом же уровне в последующие годы. По тесту на математическое и научное рассуждение (ACT тест) соотношение было 3 : 1, при этом из 19 учеников, набравших наилучшийрезультат, было18 мальчиков(Wai et al., 2010).
Гипотеза широкой нормы реакции женского пола распространяет эволюционную концепцию дифференциации полов на онтогенез, связывая в единое целое филогенетические и онтогенетические закономерности раздельнополости и взаимоотношения популяции со средой. Она позволяет объяснить феномен дифференциальной смертности полов, и многие другие непонятные онтогенетические явления, затрагивающие в разной степени мужской и женский пол, например полимерию, пенетрантность, экспрессивность, атакжепсихологическиеособенностиполов.
Приведенные рассужденияпозволяют построитьлогическуюцепочку явлений, приводящихотразной нормы реакции полов к популяционному половому диморфизму: чем шире норма реакции, тем больше онтогенетическая пластичность, тем меньше фенотипическая дисперсия и меньше смертность; чем меньше смертность, тем лучше сохраняется генотипическая дисперсия и тем меньше филогенетическая пластичность, чтоиприводитквозникновениюполовогодиморфизма.
7 8 Г Л А В А 7 . Д В А П О Л А — Д В А П О Т О К А И Н Ф О Р М А Ц И И
Глинаимрамор
Эволюция идет методом “проб и ошибок”. Чтобы локализовать их выгоднее пробовать не на целой системе, а только на её части. Для этого достаточно разделить систему на две подсистемы и обеспечить их дихронную эволюцию так, чтобы одна из них, пробная, начинала и кончала эволюцию раньше, чемосновная(болееценная).
Разделение популяции на два пола и “специализация” полов, при которой один из них “отвечает” за качество, а другой—за количество потомства, приводит к тому, что любой поток информации об изменении среды воспринимают сначала самцы, реагируют на него и преобразуют его. Такое разделение на инерционное, стабильное ядро и более изменчивую, чувствительную оболочку позволяет популяции отличить временные, коротко действующие и случайные изменения факторов среды, например необычно холодную зиму или особенно жаркое лето, от систематических изменений водномитомженаправлении, скажем, наступленииледниковогопериода.
Можно представить самцов как передовой отряд популяции, продвинутый далеко вперёд, навстречу вредным факторам среды. Между этим авангардом и “ядром”—“золотым фондом” популяции— самками выдерживается определенная дистанция, необходимая для проверки и отбора. Эволюционная инертность, отставание самок—это плата за их совершенство. И, наоборот, прогрессивность самцов—проявление их несовершенства. Таким образом, на вопрос, нужны ли самцы, можно ответить утвердительно. Они необходимы в основном для приспособления к изменившимсяусловиямсреды.
Дифференциации на женский-мужской пол, основаны на одном и том же, главном для эволюционирующих систем, принципе специализации: по сохранению (С) и изменению (И) системы. Во-первых, консервативный и оперативный аспекты эволюции—два главных ее непременных условия. Если одно из них отсутствует, то нет эволюции: система либо исчезает, либо стабильна. Во-вторых, их отношение, И/С, характеризует эволюционную пластичность системы. В- третьих, эти условия альтернативны: чем больше И, тем меньше С, и наоборот, так как они дополняют друг друга до единицы: С + И = 1. Поэтому без специализации подсистем системе приходится выбирать некий компромисный оптимум И/C, в то время, как при их специализации можно максимизировать одновременно и то и другое. В этом эволюционное преимущество всех сопряженныхдифференциаций.
* |
* |
* |
Как уже говорилось, вопрос “что дает пол?” включает в себя два самостоятельных вопроса: 1) “что дает скрещивание?” и 2) “что дает дифференциация полов?” Ответ новой теории: скрещивание обеспечивает комбинаторный потенциал, ассортимент генотипов, то есть выполняет, можно сказать генетическую функцию. А дифференциация полов обеспечивает эффективный информационный контактсосредой, изменениесреднегогенотипа, тоестьвыполняеткакбыэкологическуюфункцию.
Раздельнополое размножение выполняет две функции: генетическую—создание разнообразия генотипов путем скрещивания, и экологическую—обеспечение выгодной формы информационного контакта со средой через дифференциацию полов. Первая функция обеспечивает эволюционные преимущества половых форм размножения (гермафродитных и раздельнополых) перед бесполыми, а вторая—преимущества раздельнополых форм перед гермафродитными. Поэтому теорию можно назвать “эволюционной или генетико-экологической теорией пола”. А учитывая специализацию полов по двум главным эволюционным потокам информации: генетическому (от поколения к поколению) иэкологическому(отсреды), можноусловноназватьженскийпол“генетическим” полом, амужской—“экологическим”.
Глава8
Соотношениеполов
Третичное соотношение полов устанавливает отношение между тенденциями наследственности и изменчивости, то есть определяет эволюционную пластичность вида. Поскольку наразных этапах эволюции, а также в различных условиях среды требуется разная эволюционнаяпластичностьпопуляции, тодлякаждогоизнихсуществует своеопределенное
оптимальноезначениетретичного соотношенияполов, необязательноравное1 : 1.
Связьсоотношенияполовсусловиямисреды
В стабильных условиях среды требуется малая эволюционная пластичность, а всильно изменчивых— большая, поэтому чем стабильнее среда, тем ниже должен быть оптимум третичного соотношения полов, и, наоборот, чем изменчивее среда, тем выше. Следовательно, третичное соотношение полов является важным параметром раздельнополой популяции, определяющим ее эволюционную пластичность и тесно связанным с условиями среды. С другой стороны, среда влияет на третичное соотношение полов через дифференциальную смертность полов. Причем именно в изменчивых условиях среды повышается смертность мужского пола и падает третичное соотношение полов популяции. Следовательно, мы опять имеем дело с конфликтной ситуацией: чем изменчивее среда, тем меньше остается в популяции мужских особей, и одновременно, тем больше их требуется для приспособления, тоестьвизменчивыхусловияхпопуляцияможетвообщеутратитьмужскойпол.
Единственная возможность избежать потери мужских особей—наличие механизма, позволяющего сохранятьили даже повышать третичное соотношение половвэкстремальных условиях. Этого можно добиться только путем повышения вторичного соотношения полов в таких условиях. Следовательно, во всех экстремальных условиях среды, когда понижается третичное соотношение полов, одновременно должно повышаться вторичное, и наоборот, в оптимальных условиях, когда повышается третичное соотношение полов, должно понижаться вторичное. Иными словами, в экстремальных условиях среды, будет одновременно повышаться и смертность и рождаемость мужских особей, то есть будет расти их “оборачиваемость”. Следовательно, вопреки существующему взгляду на вторичное соотношение полов как на константу, характерную для данного вида, которая обеспечивает оптимум третичного соотношения полов 1 : 1, согласно новой концепции, вторичное соотношение полов является переменной величиной, тесно связанной с условиями среды. В стабильных, оптимальных условиях, когда не требуется много мужских особей, вторичное соотношение полов должно понижаться. В изменчивых, экстремальных условиях, когда требуется многомужских особей, наоборот, онодолжно повышаться (Геодакян, 1978).
79