

Механизм действия гормонов, взаимодействующих с 1-тмс-рецепторами
К 1-TMС-(R) относятся гуанилатциклазы, катализирующие образование цГМФ из ГТФ. Первый тип гуанилатциклаз, локализующийся в плазматической мембране, активируется внеклеточными лигандами (предсердным натрийуретическим фактором); второй —цитозольный фермент, активируемый оксидом азота (NO۬). Оксид азота вызывает расслабление гладкой мускулатуры сосудов, в том числе коронарных.
Гормон, присоединившись к 1-TMС-(R), его активирует. Этот гормон-рецепторный комплекс проявляет гуанилатциклазную активность, катализируя образование цГМФ из ГТФ. цГМФ (вторичный посредник) активирует протеинкиназу С, которая начинает фосфорилировать остатки Сер и Тре в белках-мишенях, что приводит эти белки в функционально активное состояние. цГМФ является также активатором фосфодиэстераз, катализирующих распад цАМФ (т. е. цГМФ и цАМФ – антагонисты). Конечный эффект цГМФ проявляется в изменении транспорта ионов и воды в почках и кишечнике; в сердечной мышце цГМФ способствует мышечному расслаблению; в палочках и колбочках сетчатки глаза цГМФ стимулирует открытие ионных каналов.
Самой распространённой группой 1-TMС-(R) являются рецепторные тирозинкиназы. В их структуре имеется 4 домена: внеклеточный (связывает гормон), трансмембранный, внутриклеточный с тирозинкиназной активностью, внутриклеточный регуляторный. Важнейшей тирозинкиназой является (R) инсулина. Присоединение инсулина вызывает аутофосфорилирование киназного домена (R). При этом ускоряется фосфорилирование остатков тирозина в особом белке — IRS-1 (инсулин-рецепторный субстрат). Активный IRS-1 включает целый каскад реакций, усиливающих гормональный сигнал. Это протеинкиназные реакции с участием, в первую очередь, МАП-киназ (митоген-активируемые протеинкиназы). МАП-киназы изменяют активность многих ферментов путём их фосфорилирования.
В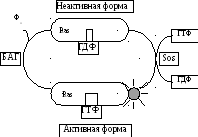 передаче сигнала с участием1-TMС-(R)
есть свои выключатели.
ЭтоRas-белки, относящиеся к семейству
мономерных ГТФ-аз. Ras-белки могут быть
активными, если к ним присоединяется
ГТФ, и становиться неактивными при
замене ГТФ на ГДФ. Однако Ras-белки
катализируют гидролиз ГТФ очень медленно.
Существует специальный белок, ускоряющий
гидролиз ГТФ, —БАГ(белок,
активирующийГТФ-азу). Другой
специальный белок —SOS-белок(фактор, высвобождающий гуаниловые
нуклеотиды) — препятствует этому
гидролизу, заменяя ГДФ на ГТФ, поэтому
он оказывает активирующий эффект наRas-белок.
передаче сигнала с участием1-TMС-(R)
есть свои выключатели.
ЭтоRas-белки, относящиеся к семейству
мономерных ГТФ-аз. Ras-белки могут быть
активными, если к ним присоединяется
ГТФ, и становиться неактивными при
замене ГТФ на ГДФ. Однако Ras-белки
катализируют гидролиз ГТФ очень медленно.
Существует специальный белок, ускоряющий
гидролиз ГТФ, —БАГ(белок,
активирующийГТФ-азу). Другой
специальный белок —SOS-белок(фактор, высвобождающий гуаниловые
нуклеотиды) — препятствует этому
гидролизу, заменяя ГДФ на ГТФ, поэтому
он оказывает активирующий эффект наRas-белок.
Механизм действия гормонов, взаимодействующих с внутриклеточными (r)
Витамин Д, ретиноевая кислота, стероидные и тиреоидные гормоны взаимодействуют непосредственно с внутриклеточными (R), поскольку они, будучи гидрофобными, свободно диффундируют через плазматическую мембрану. Эти рецепторы делятся на: 1) (R) стероидных гормонов (минералокортикоидов, андрогенов, прогестерона); 2) (R) тиреоидных гормонов (в том числе витамина Д и ретиноевой кислоты).
(R) стероидных гормонов находятся в цитозоле. Они образуют комплексы с белками теплового шока hsp (heat shock proteins), которые присоединяются к (R) таким образом, что закрывают его ДНК-связывающий домен. Кода гормон взаимодействует с (R), то белок-ингибитор hsp отделяется от (R), в результате чего (R) активируется. Активированный гормон-рецепторный комплекс перемещается в ядро, где он взаимодействует со строго определенными гормон-узнающими элементами ДНК (hormone response element).
(R) тиреоидных гормонов находятся в ядре в связанном с хроматином состоянии (за исключением рецепторов эстрогенов, которые в ядре образуют комплексы с hsp). После взаимодействия с гормоном изменяется конформация (R), что повышает его сродство к ДНК, т. е. (R) может связываться со специфическими генами в ядре, регулируя их экспрессию.