

Гормоны. Общий механизм действия гормонов
Гормоны — это класс регуляторных молекул, синтезируемых специальными клетками.
Особенности биологического действия: 1) низкая концентрация в крови (10–6–10–12 М); 2) обязательная связь с рецептором – (R), включающим каскадный механизм усиления гормонального сигнала; 3) изменение скорости синтеза ферментов или их активности; 4) регуляция секреции по принципу прямой и (или) обратной связи.
Взаимодействие гормона и (R) характеризуется высокой специфичностью, которая обеспечивается комплементарностью между структурой гормона и активного центра (R). В результате эффекта кооперативности, возникающего при взаимодействии гормона и (R), существенно изменяется активность (R) — это есть феномен амплификации (усиления) гормонального сигнала. Механизм амплификации включает участие специальных ферментов и молекул — вторичных посредников. Гормональный сигнал способен «выключаться» в результате инактивирования (R) путём его фосфорилирования, либо удаления (R) с поверхности клетки (эндоцитоз) и т. д. Множество разных сигналов, воспринимаемых клеткой, суммируется в один определённый ответ.
Клеточные (R) в зависимости от их локализации делятся на 2 большие группы: 1) (R) плазматической мембраны; и 2) внутриклеточные (R).
(R) плазматической мембраны клеток обеспечивают узнавание, связывание и передачу регуляторного сигнала внутрь клетки. Среди них различают:
1. 7-ТМС-(R) — это интегральные мембранные белки с семью трансмембранными спиральными сегментами, соединенными гидрофильными внеклеточными и внутриклеточными петлями. Внутриклеточные петли этих (R) содержат центры связывания G-белка.
2. 1-TMС-(R) — это интегральные мембранные белки с одним трансмембранным сегментом и глобулярными доменами на вне- и внутриклеточной поверхностях мембраны. Внеклеточный домен содержит участок узнавания и связывания гормона, а внутриклеточный обладает каталитической активностью. Когда (R) активируется гормоном, его внутриклеточный домен катализирует образование внутриклеточных вторичных посредников.
3. Каналообразующие (R) — состоят из белковых субъединиц, каждая из которых содержит несколько трансмембранных сегментов.
Внутриклеточные (R) расположены в цитозоле или ядре клетки. После связывания с гормоном они изменяют скорость транскрипции и трансляции определённых генов.
По химической природе гормоны делят на:
пептиды (глюкагон, котрикотропин) и белки (сложные белки — тиреотропин, гонадотропины; простые белки — соматотропин, инсулин);
производные аминокислот (адреналин, серотонин, тироксин);
стероиды (альдостерон, кортизол, половые гормоны, витамин Д и ретиноевая кислота);
производные липидов (эйкозаноиды).
Механизм действия гормонов, взаимодействующих
с 7-ТМС-рецепторами
Первый этап действия гормона заключается во взаимодействии гормона с 7-ТМС-(R) (к ним относятся α- и β-адренергические (R), (R) гистамина, серотонина, соматостатина, гликопротеиновых гормонов, глюкагона, паратирина, кальцитонина, гормонов гипоталамуса).
 Второй
участник передачи гормонального сигнала
—G-белки.
Обнаружены: 1) большие, состоящие из
нескольких субъединиц G-белки
(связаны с мембранами); 2) низкомолекулярные,
состоящие из одной полипептидной цепи
(цитозольные). Все G-белки
обладают ГТФ-азной активностью, и их
конформация зависит от того, связаны
ли они в данный момент с ГДФ или ГТФ.
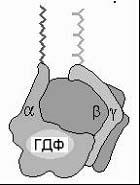
Мембранный G-белок
состоит из 3 субъединиц: α, β и γ.
Второй
участник передачи гормонального сигнала
—G-белки.
Обнаружены: 1) большие, состоящие из
нескольких субъединиц G-белки
(связаны с мембранами); 2) низкомолекулярные,
состоящие из одной полипептидной цепи
(цитозольные). Все G-белки
обладают ГТФ-азной активностью, и их
конформация зависит от того, связаны
ли они в данный момент с ГДФ или ГТФ.
Мембранный G-белок
состоит из 3 субъединиц: α, β и γ.
В неактивном состоянии три субъединицы соединены вместе, и α-субъединица связана с ГДФ. После присоединения гормона к 7-ТМС-(R) и взаимодействия гормон-рецепторного комплекса с G-белком в последнем происходит замена ГДФ на ГТФ, вследствие чего G-белок диссоциирует с образованием свободной α-субъединицы и димера субъединиц β, γ. Затем α-субъединица перемещается по мембране и взаимодействует с мембраносвязанными ферментами (аденилатциклазой или фосфолипазой С), катализирующими образование низкомолекулярных вторичных посредников. Продолжительность эффекта G-белка определяется ГТФ-азной активностью его α-субъединицы: после гидролиза ГТФ до ГДФ действие субъединицы прекращается и G-белок возвращается в исходное тримерное состояние
Аденилатциклаза. Различные типы аденилатциклаз реагируют с α-субъединицами, выполняющими разные функции: если связывание осуществляется с αs-субъединицей G-белка, то фермент активируется, если с αI-cубъединицей, то ингибируется. Аденилатциклаза катализирует образование цАМФ из АТФ. Уровень цАМФ в клетках очень низок, но при активировании аденилатциклазы он может быстро и значительно увеличиваться. Однако это повышение кратковременно. Распад цАМФ катализируется цАМФ-фосфодиэстеразами (ФДЭ), гидролизующими фосфодиэфирную связь в молекуле цАМФ. ФДЭ — регулируемый фермент (его ингибирует кофеин, что приводит к повышению уровня цАМФ).
ц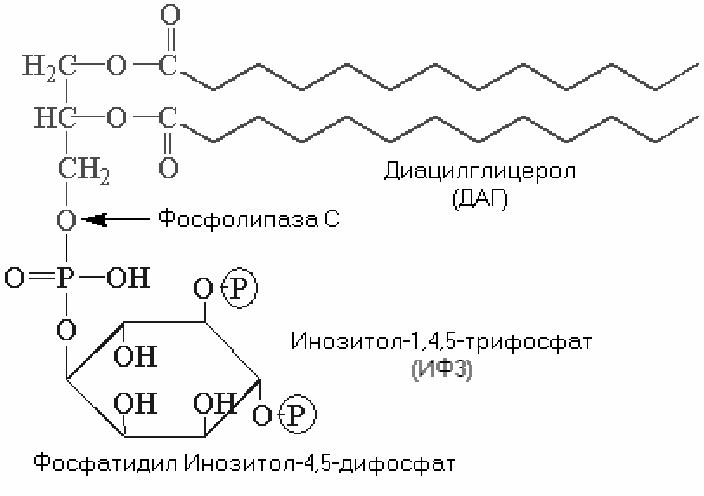 АМФ
является аллостерическим регуляторомпротеинкиназы
А (существуют
разные типы этих ферментов).
Протеинкиназы
— это гетеромеры, состоящие из
двухкаталитических и одной регуляторной
субъединиц. Присоединение цАМФ к
регуляторной субъединице высвобождает
каталитические субъединиц. Последние
переносят фосфорный остаток с АТФ на
серин или треонин некоторых ферментов,
которые при этом переходят в активное
или неактивное состояние (это зависит
от свойств данного фермента: так,
фосфорилирование
гликогенфосфорилазы
активирует ее, а фосфорилирование
гликогенсинтазы её угнетает). цАМФ
регулирует также открытие цАМФ-зависимых
ионных каналов. Повышение уровня цАМФ
может приводить к активации транскрипции
определенных генов.
АМФ
является аллостерическим регуляторомпротеинкиназы
А (существуют
разные типы этих ферментов).
Протеинкиназы
— это гетеромеры, состоящие из
двухкаталитических и одной регуляторной
субъединиц. Присоединение цАМФ к
регуляторной субъединице высвобождает
каталитические субъединиц. Последние
переносят фосфорный остаток с АТФ на
серин или треонин некоторых ферментов,
которые при этом переходят в активное
или неактивное состояние (это зависит
от свойств данного фермента: так,
фосфорилирование
гликогенфосфорилазы
активирует ее, а фосфорилирование
гликогенсинтазы её угнетает). цАМФ
регулирует также открытие цАМФ-зависимых
ионных каналов. Повышение уровня цАМФ
может приводить к активации транскрипции
определенных генов.
Фосфолипаза С. Субстратом этого фермента является фосфатидилинозитол-4,5-дифосфат. Фосфолипаза С катализирует образование двух мощных вторичных посредников: диацилглицерола (ДАГ) и инозитол-1,4,5-трифосфата (ИТФ).
ИТФ связывается с (R) кальцисом, что приводит к открытию лиганд-зависимых кальциевых каналов кальцисом. В результате в цитозоле резко увеличивается концентрация Са2+. Высвободившийся Са2+ захватывается специальными кальций-связывающими белками, например, калмодулином. Когда уровень внутриклеточного Са2+ повышается, кальций-калмодулиновый комплекс присоединяется к различным белкам-мишеням, что сопровождается изменением их активности. Кальмодулин-зависимые протеинкиназы регулируют активность многих ферментов. Они также способны проникать в ядро клетки, где фосфорилируют белки, ответственные за активацию транскрипции.
Таким образом, ИТФ повышает концентрацию Са2+ в цитозоле. ДАГ остается в плазматической мембране и либо гидролизуется с образованием арахидоновой кислоты (необходимой для синтеза простагландинов), либо активирует протеинкиназу С. При низком внутриклеточном уровне Са2+ и отсутствии ДАГ протеинкиназа С находится в цитоплазме в неактивном состоянии. При связывании ДАГ конформация протеинкиназы С изменяется, увеличивается её сродство к Са2+ и она начинает активировать разнообразные белки-мишени (так, С-киназа фосфорилирует и тем самым активирует Na+/H+ обменный насос плазматической мембраны, контролирующий внутриклеточный рН). Активная С-киназа способна усиливать транскрипцию генов.