

13. Поверхностный аппарат эукариотической клетки. Строение и функции.
См. 10 вопрос.
14. Транспортная роль белков плазмалеммы.
См.10 и 11 вопросы.
15. Активный транспорт веществ через мембрану. К+- Nа+ насос.
См. 11 вопрос
16. Поверхностный аппарат клетки. Транспорт макромолекул.
См. 12 (то же самое)
17. Рецепторная роль плазмалеммы.
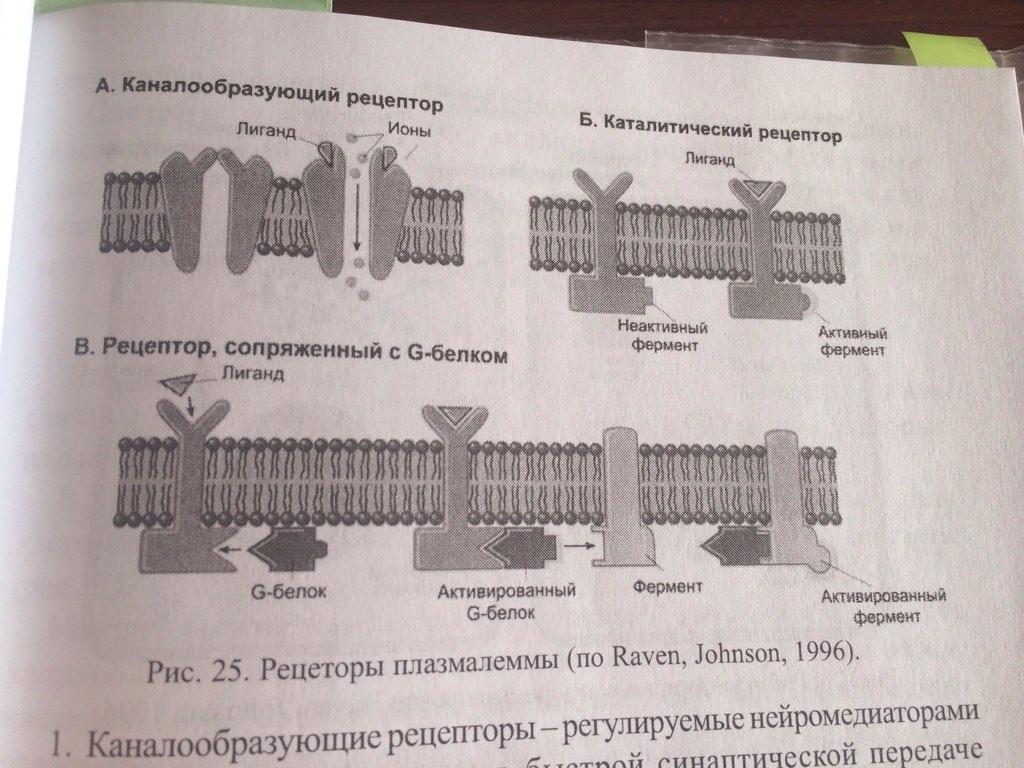
Плазмалемма, биологическая мембрана, содержит различные сигнальные белки. Все водорастворимые и некоторые жирорастворимые сигнальные молекулы присоединяются к спецефическим белковым рецепторам на поверхности клеток-мишений. В зависимости от способа перечади сигнала внутрь клетки рецепторы можно разделить на три класса. 1. Каналообразующие рецепторы-регулируемые нейромедиаторами ионные каналы, участвующие в быстрой синаптической передачи импульсма. 2-каталитические рецепторы- при активации лигандом начинают работать как ферменты, фосфорилирую определенные белки.(напр, рецепторы инсулина) 3. Рецепторы, сопряженные с G-белками. Активация этих рецепторов запускает целую цепь реакций, изменяющих концентраци. В цитоплазме малых сигнальных молекул, которые называются вторичными посредниками. Наиболее важными вторичными посредниками являются циклические АМФ и Са. Эти молекулы могут влиять на поведение белков-мишеней в клетке, активирую обмен веществ. Аденилатциклазная система:мембрана скелетных мыщц, жировые клетки, клетки сердцечной мыщцы. Сигнальными молекулами для этой системы служат многие гормоны и локальные химические медиаторы. Когда соответствующий лиганд ( например, адреналин) взаимодействует с рецептором, происходят конфармационные изменения рецепторного белка. Это вызывает структурную перестройку G-белка, находящегося в мембране. G-белок в этом состоянии активизирует адениатциклазу-фермент, также распологающийся в мембране. Активизированнная аденилатциклаща катализирует превращение АТФ в ц-АМФ. Ц-Амф воздействует на ферменты протеинкиназы, которые участвуют во многих метаболических процессах в клетке.
ИЛИ
ЦИТОРЕЦЕПТОРЫ
Взаимодействия клетки с её окружением осуществляется при участии специальных структур, локализованных на плазмолемме –
рецепторов. Существуют рецепторы к биологически активным веществам — гормонам, медиаторам, к специфическим антигенам
разных клеток или к определенным белкам. Рассматривая клетку как ячейку биологических организмов, снабженную рецепторами, необходимыми для поддержания постоянства ее внутренней среды и обеспечивающими адекватное реагирование на изменение внешних условий, становится очевидным, что рецепторы располагаются на цитоплазматической мембране. Рецепторами на поверхности клетки могут служить гликопротеиды и гликолипиды мембран. Считается, что такие чувствительные к отдельным веществам участки могут быть разбросаны по всей поверхности клетки или собраны в небольшие зоны. Особый вид поверхностных рецепторов – рецепторы, связанные с G-белками. Это трансмембранные белки, которые могут быть связаны либо с ионным каналом, либо с ферментом. Состоят из двух частей: рецепторной, взаимодействующей с сигнальными молекулами, и субъединиц G-белка α, β, γ. G-белки – белки, связывающие гуанозинтрифосфат (ГТФ). После связывания с сигнальной молекулой комплекс G-белков передает сигнал на ассоциированный с цитолеммой фермент аденилатциклазу, которая синтезирует вторичный посредник циклический аденозинмонофосфат (цАМФ). В качестве вторичного посредника могут выступать и молекулы кальция. Через рецепторы, связанные G- белками, опосредуется действие на клетку подавляющего большинства гормонов и нейромедиаторов. Внутриклеточные рецепторы находятся внутри клетки – в гиалоплазме, на мембранах органелл (цитоплазматические рецепторы), в ядре (ядерные рецепторы). Они предназначены для гормонов и других биологически активных веществ, которые в силу неполярности своих молекул могут легко проникать внутрь клетки
(стероидные и тиреоидные гормоны и др.). Особый интерес представляют ядерные рецепторы. С этими рецепторами связываются такие гормоны, как стероидные, тиреоидные, витамин D3. Молекулы таких рецепторов состоят из 2 участков: участок для связывания с гормоном и участок, взаимодействующий соспецифическими участками ДНК в ядре. Ядерные рецепторы являются факторами транскрипции. Некоторые из них относятся к протоонкогенам – генам нормального генома, регулирующим пролиферацию клеток органов-мишеней, их дифференцировку и межклеточные взаимодействия. В результате соматических мутаций в протоонкогенах может происходить злокачественное перерождениеклеток.
18. Особенности воздействия на клетки-мишени гидрофильных и гидрофобных сигнальных молекул.
Реакция клетки на внеклеточный сигнал осуществляется клетками-мишенями с помощью белков-рецепторов, которые связывают сигнальную молекулу и стимулируют ответную реакцию. Сигнальные молекулы, в зависимости от своей растворимости, связываются с поверхностными или внутриклеточными рецепторами. Жирорастворимые молекулы- стероидные гормоны проходят через плазматическую мембрану и активируют рецепторный белок внутри клетки. Водорастворимые молекулы- большинство гормонов и нейромедиаторы, активируют рецептор на поверхности клеток.
Все водорастворимые и некоторые жирорастворимые сигнальные молекулы присоединяются к специфическим белковым рецепторам на поверхности клеток-мишеней. В зависимости от способа передачи сигнала рецепторы делятся на три класса:
Каналообразующие рецепторы-регулируемые нейромедиаторами ионные каналы, участвующие в быстрой синаптической передаче импульса.
Каталитические рецепторы - при активации лигандом начинают работать как ферменты, фосфорилируя определенные белки. К этой I руппе относятся рецепторы инсулина и многих ростовых факторов.
Рецепторы, сопряженные с G-белками, кот наз вторичными посредниками. Наиболее важные вторичные посредники-циклический АМФ ( ц-АМФ ) и Са2+, кот могут влиять на поведение белков-мишеней, активируя обмен вещ.
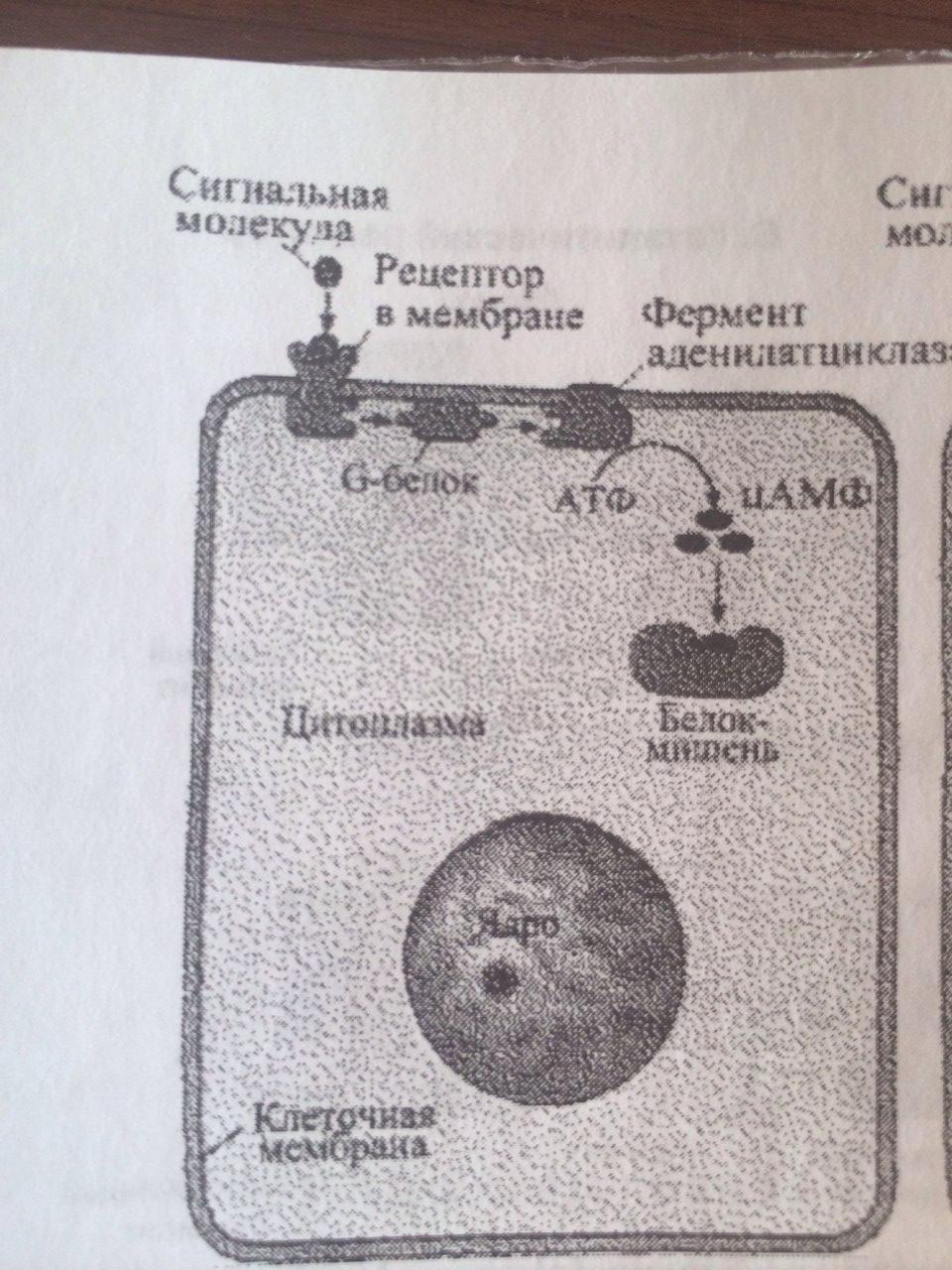
Способы передачи сигнала в клетки-мишени от сигнальных молекул: 1. АДЕНИЛАТЦИКЛАЗНАЯ (ИЛИ ГУАНИЛАТЦИКЛАЗНАЯ) СИСТЕМА (в мембр скел мышц, жир кл и др) Сигнальными молекулами для этой системы служат многие гормоны и локальные химические медиаторы. Когда соответствующий лиганд (например, адреналин) взаимодействует с рецептором, происходяi конформационные изменения рецепторного белка. Это вызывао структурную перестройку G-белка, находящегося в мембране. G-белок в этом состоянии активизирует аденилатциклазу - фермент, также располагающийся в мембране. Активизированная аденилатциклаза катализирует превращение АТФ в ц-АМФ. Ц-АМФ воздействует на ферменты протеинкиназы, которые участвуют во многих метаболических процессах в клетке.
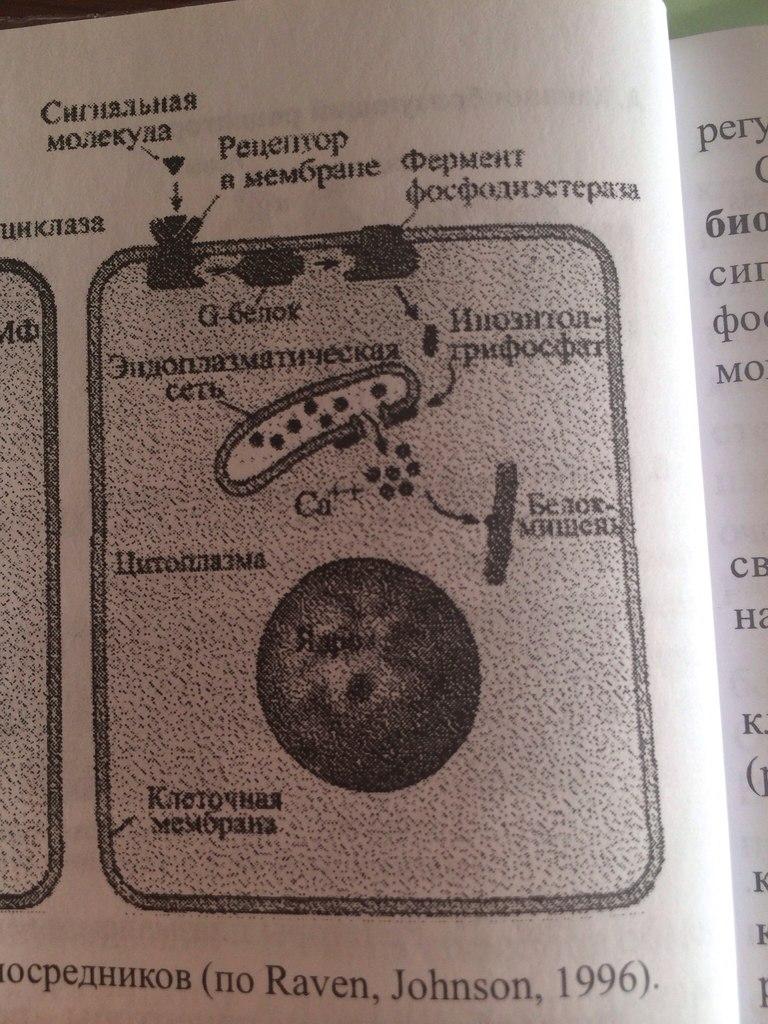
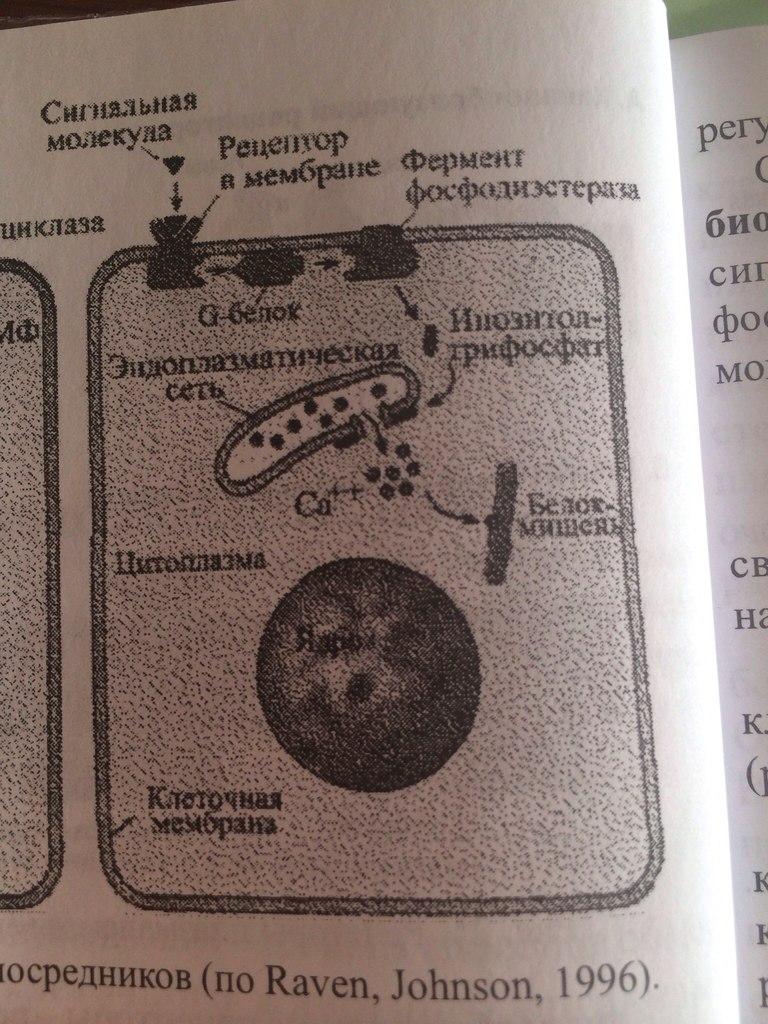
2. ФОСФОИНОЗИТИДНЫЙ МЕХАНИЗМ. Внутриклеточная система инозитолтрифосфата (ИФ3) имеет сходство с системой ц-АМФ. Как и в системе цАМФ, внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са2+ из эндоплазматической сети. Повышение концентрации Са2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты.
19.Сигнализация с участием рецепторов клеточной поверхности и вторичных посредников на примере ц-AMФ.
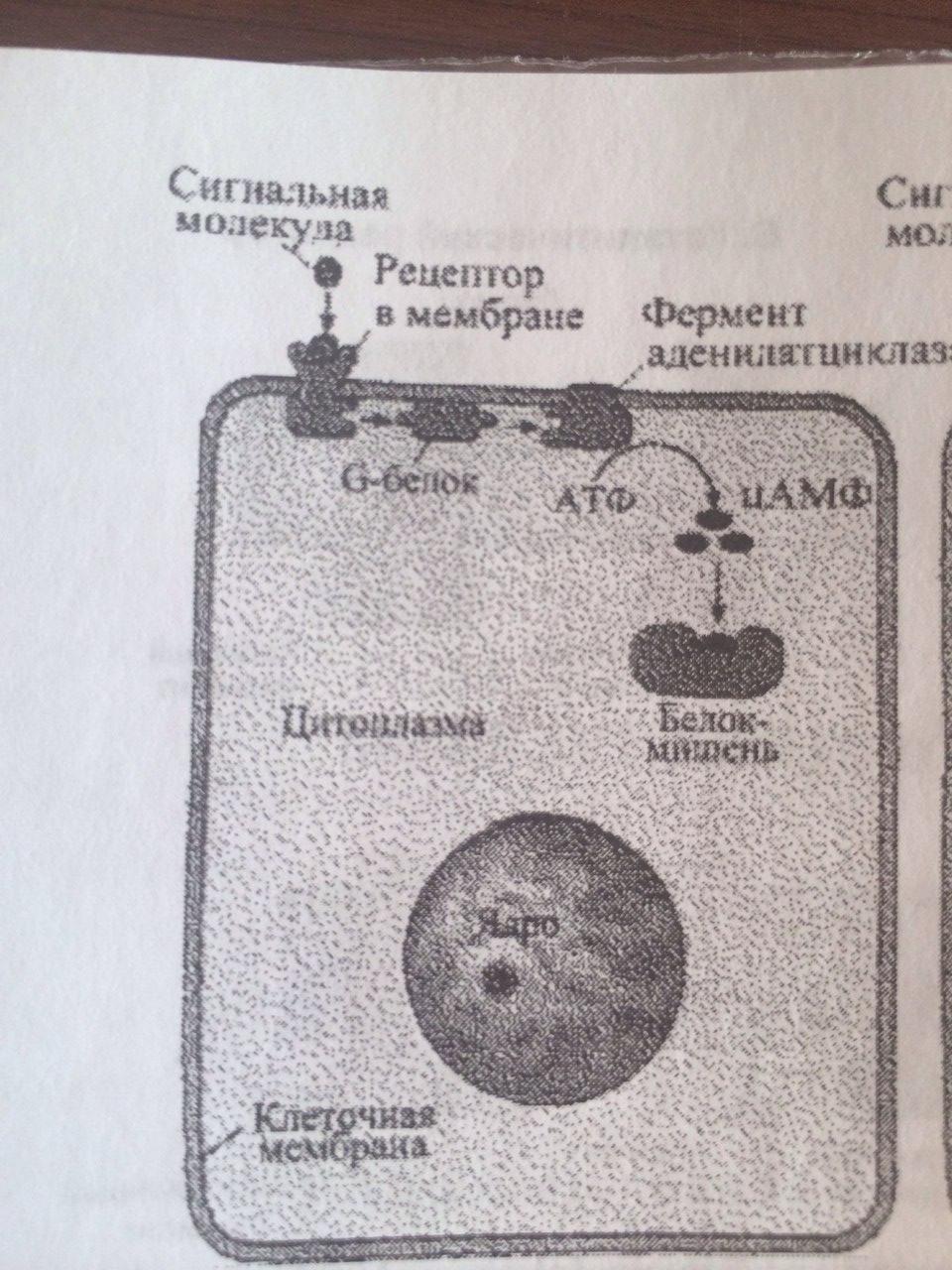
В системе ц-АМФ, внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са2+ из эндоплазматической сети. Повышение концентрации Са2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты, участвующ во многих метаболических процессах в клетке.
20. Системы сигнализации: Эндокринная, Синаптическая. Роль медиаторов и гормонов.
Сигнал передается через жидкие среды организма (кровь, лимфу, межклеточную жидкость). Это гуморальная система связи - эндокринная сигнализация.
При получении сигнала клетки изменяют режим функционирования. Эндокринные клетки обычно собраны в специальные железы и выделяют гормоны во внеклеточную жидкость, окружающую все клетки в тканях. Отсюда гормоны проникают в капилляры, в кровоток, опять в капилляры, межклеточную жидкость и улавливаются клетками "мишенями".
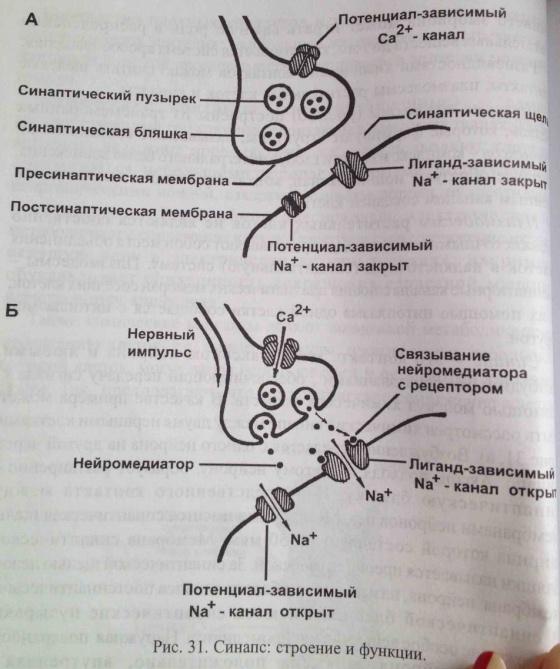
Эндокринная сигнализация не может быть использована для передачи сигнала нервной системы. Здесь сигнал должен быть очень краток во времени и конкретен по адресу передачи. Работа нервных клеток отличается гораздо большей быстротой и точностью. Они передают информацию на большие расстояния по нервному волокну с помощью электрических импульсов с высокой скоростью. Только в нервных окончаниях импульсы преобразуются в химические сигналы. В нервном окончании высвобождается нейромедиатор. Он улавливается постсинаптической мембраной в следующем нейроне и стимулирует выработку электрического сигнала. Такая передача сигнала называется синаптической. Синапсисом называется щель между двумя соседними нервными клетками. Она используется только в нервной системе. В отличие от гормонов, нейромедиаторы действуют только на постсинаптическую мембрану. Они улавливаются специальными белками - рецепторами. Рецепторы связывают сигнальную молекулу и инициируют ответ.
СИНАПС. Когда нервный импульс достигает окончания аксона, происходит изменение мембранного потенциала-деполяризия мембраны аксона. Деполяризация вызывает временное открываение кальциевых каналов в мембране. Посколько концентрация Ca снаружи превышает концентрацию свободного Ca в клетке, Ca устремляется внутрь нервного окончания. В результате этого, синаптический пузырьки сливаются с пресимпатической мембраной и нейромедиатор путем экзоцитоза выводится в синаптическую щель. Молекулы нейромедиатора диффундируют через щель, связываясь с рецепторами. Каналы рецепторов открываются, посколько концентрация ионов Na cнаружи клетки выше, чем внутри, ионы nA устремляются внутрь нейрона. В результате-деполяризация мембраны
21.Понятие о вторичных посредниках. Инозитолфосфатная система.
Вторичные (внутриклеточные) мессенджеры – малые молек или ионы, внутриклет концентрац кот повышает ответ на соединение рецептора с сигнальной молекулой. Роль вторичных мессенджеров выполняют молекулы и ионы: • ц-АМФ (циклический аденозин-3',5'-монофосфат); • ИФ3 (инозитол-1,4,5-трифосфат); • ДАГ • ц-ГМФ • Са2+.
Внутриклеточная система инозитолтрифосфата (ИФ3) имеет сходство с системой ц-АМФ. Как и в системе цАМФ, внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са2+ из эндоплазматической сети. Повышение концентрации Са2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты.
22.Понятие о вторичных посредниках. Аденилатциклазная система.
Вторичные (внутриклеточные) мессенджеры – малые молек или ионы, внутриклет концентрац кот повышает ответ на соединение рецептора с сигнальной молекулой. Роль вторичных мессенджеров выполняют молекулы и ионы: • ц-АМФ (циклический аденозин-3',5'-монофосфат); • ИФ3 (инозитол-1,4,5-трифосфат); •ДАГ •ц-ГМФ • Са2+.
Аденилатциклазная система включает: - интегральные белки цитоплазматической мембраны: • Rs - рецептор первичного мессенджера – активатора, рецептор первичного мессенджера - ингибитора АЦС; • фермент аденилатциклазу (АЦ). - «заякоренные» белки: • Gs - ГТФ-связывающие белки • цитозольный фермент протеинкиназу А (ПКА).
Рис. 4.11. Функционирование аденилатциклазной системы

|
|
Когда соответствующий лиганд (например, адреналин) взаимодействует с рецептором, происходяi конформационные изменения рецепторного белка. Структурно перестроенный G-белок активизирует аденилатциклазу - фермент, располагающийся в мембране. Активизированная аденилатциклаза катализирует превращение АТФ в ц-АМФ. В системе ц-АМФ, внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са2+ из эндоплазматической сети. Повышение концентрации Са2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты, участвующ во многих метаболических процессах в клетке. Участие аденилатциклазной системы в регуляции экспрессии генов. Многие белковые гормоны: глюкагон, вазопрессин, паратгормон и др. могут не только изменять скорости реакций путем фосфорилирования ферментов, но и увеличивать или уменьшать их количество, регулируя экспрессию генов.
23. Принципы и механизмы преобразования сигналов в клетке.
-Изменения конформации белков -Изменение активности путем фосфо и дефосфорилирования белков (фосф к-ты из АТФ) -Преобразование сигнала с уч. G-белков -Обравзование вторичных посрелников Ферменты: 1)Киназы-фосфорилировае. 2)Фосфотазы-дефосфорилирование Каталитические рецепторы. Белки-рецепторы.(Инсулин, и многие фкторы роста) У кот домен облад ферментативной(киназной)СПОСОБНОСТЬЮ АКТИВАЦИИ БЕЛКОВ. G-белки – разновидность GTPазы, стр и активность кот зависит от нахождения ГТФ или ГДФ-связном состоянии. Участвуют в передаче неклеточных сигналов от рецепторов в клетку (сопрягая активацию рецепторов в работой ионных каналов или ферментов) РОЛЬ G-белков: (Гилман и Мартин-открытие) •Усиление сигнала •Возможность регудяции активности белков-мишений.(G5-стимулирующ.G1-ингибирующ белок) (Активац калиевого канала с уч G-белка) В системе ц-АМФ(Аденилатциклазная), внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са2+ из эндоплазматической сети. Повышение концентрации Са2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты, участвующ во многих метаболических процессах в клетке. Участие аденилатциклазной системы в регуляции экспрессии генов. Многие белковые гормоны: глюкагон, вазопрессин, паратгормон и др. могут не только изменять скорости реакций путем фосфорилирования ферментов, но и