

Позиционные сигналы и пролиферация клеток
(продолжение)
Механизм этих сдвигов инициируется транслокацией нефосфорилированного β-катенина в ядро, где он взаимодействует с ядерным фактором транскрипции TCF/LEF-1. Образующийся комплекс β-катенин - TCF воздействует на промоторный участок гена циклина D1, что приводит к экспрессии его транскрипции и, как следствие этого, - к пролиферации клеток. Одновременно LEF-1 воздействует на ген кадгерина Е, который является для него мишенью. Под влиянием LEF-1 происходит супрессия данного гена, в результате чего снижается синтез катгерина Е и ослабляется межклеточная адгезия. Соответственно, уменьшается эффект "контактного торможения", что способствует клеточной пролиферации, повышению инвазивности и эмиграции клеток.
Уменьшение экспрессии кадгерина Е сопровождается снижением экспрессии и интегринов. В результате ослабляется взаимодействие клеток с внеклеточным матриксом, что также увеличивает клеточную подвижность.
Таким образом, Е-кадгерин-катениновый комплекс, участвующий в формировании межклеточных контактов функционирует и как супрессор опухоли, а экспрессирующие его гены могут рассматриваться как гены - супрессоры. Ослабление экспрессии этого комплекса или его несостоятельность, обусловленная мутациями в гене кадгерина Е (наблюдаемые в половине случаев рака желудка и при раке яичников у человека), а также мутациями и делециями в генах, кодирующих α и β-катенины служит позиционным сигналом к клеточной пролиферации и важнейшим условием перехода опухоли к инвазивному фенотипу. С этими представлениями связано определение экспрессии Е-кадгерина у онкологических больных в целях прогнозирования рецидивов, а восстановление нормальных адгезивных функций кадгерин-катенинового комплекса является задачей антиинвазивной терапии опухолей.
Позиционный сигнал к пролиферации клеток может исходить и от других молекул адгезии - интегринов, обеспечивающих контакт "клетка-матрикс". С подмембранным доменом этих молекул (интегрины 1 и 3) связана интегринассоциированная киназа - ILK. Это серин-треониновая киназа, непосредственно взаимодействует с GSK-3, фосфорилирует ее и ингибирует тем самым ее активность. Таким образом, активация ILK блокирует перевод β-катенина в фосфорилированную форму, что определяет возможность его ядерной транслокации.
Блокада GSK-3, стимулирующая клеточную пролиферацию, может осуществляться при участии Wnt-сигнального пути, который играет чрезвычайно важную роль в эмбриогенезе, в частности, в эмбриональной индукции, в формировании полярности эмбриональных клеток и в определении их дальнейшей судьбы. Этот путь, начинающийся со взаимодействия Wnt со своим рецептором Frizzied c помощью белков Dishevellid и аксина также замыкается на GSK-3, ингибирует ее активность, способствуя проникновению нефосфорилированного β-катенина в ядро.
Позиционные сигналы и пролиферация клеток
(по Ю.А. Ровенскому, 1998; Е.Д. Свердлову, 2001)

Внеклеточный
матрикс



Cа2+
Cа2+ α β
Frizzied
Cа2+
Cа2+ Кадгерин Интегрин


САРКОЛЕММА
Dichevelled









Аксин






Gsk-3 Р Р



P







Деградация














Актин Ядерная
мембрана







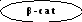







Транслокация
Ген циклина D1
Экспрессия
транскрипции циклина D1

Ген
кадгерина Е


Супрессия
транскрипции кадгерина Е
Пролиферация
клетки





Снижение
синтеза кадгерина
Е
Ослабление
межклеточной адгезии Ослабление
контактного торможения Инвазивность
и эмиграция
клеток



Снижение
экспрессии кадгерина Е на клеточ-ной
поверхности
Снижение
экспрессии интегринов
Ослабление
взаимодей-ствия клеток с внекле-точным
матриксом
Увеличение
клеточной подвижности




Раздел III.
ПРОТООНКОГЕНЫ. ОНКОГЕНЫ. ОНКОСУПРЕССОРЫ.
ОПРЕДЕЛЕНИЕ ПОНЯТИЙ
(по Б.П. Копнину, 2000; А.В. Лихтенштейну, В.С. Шапоту, 2001)
Онкогены - гены (последовательности нуклеиновых кислот), обусловливающие неконтролируемый опухолевый рост клеток (in vivo) и их трансформацию в культуре (in vitro). Выделяют вирусные (v-onc) и клеточные (c-onc) онкогены.
Вирусные онкогены - трансформирующие гены в составе высокоонкогенных РНК и ДНК-содержащих вирусов.
Онкогены ДНК-содержащих опухолеродных вирусов вирусспецифичны и клеточных аналогов не имеют.
Онкогены РНК-содержащих опухолеродных вирусов (ретровирусов) - исходно не вирусного происхождения и не нужны для размножения вирусов. Они происходят из протоонкогенов, захваченных вирусом из генома позвоночных во время инфекционного цикла.
Протоонкогены - клеточные гомологи вирусных (ретровирусы) онкогенов в геноме позвоночных. Это - нормальные гены, контролирующие синтез факторов роста, белков-рецепторов, белков-трансдукторов митогенного сигнала, факторов транскрипции, а через них - процессы клеточной пролиферации, дифференцировки и морфогенеза. Протоонкогены - элементы позитивной регуляции клеточного деления. Они являются его акселераторами.
Не все протоонкогены, обнаруженные в геноме человека, имеют вирусные аналоги.
В нормальной клетке протоонкогены характеризуются адекватным функционированием в соответствии с поступающими сигналами, определяющими потребность клетки в контролируемых ими белках. Если протоонкогены переходят в перманентно активное состояние вне зависимости от поступающих сигналов, они вызывают неконтролируемый рост и трансформацию клетки. В этом случае их называют клеточными онкогенами (с-onc).
Активированные с-onc независимы от клеточного генома. Они инициируют и поддерживают размножение трансформированных клеток, активируя сигнальные механизмы митотического цикла.
Структура онкогенов ретровирусов отличается от структуры протоонкогенов. Большинство из этих структурных изменений обусловливает возможность v-onc встраиваться в хромосомы хозяина и индуцировать активацию клеточных протоонкогенов, превращая их в c-onc.