

Роль макрофагов в противоопухолевой защите
LFA-1 АТ
к СТОА МФ + МФ






Опухолевая
клетка-мишень ICAM-1
Опухолевая
клетка-мишень, экспрессирующая СТОА СТОА МФ
Прикрепление
МФ к клетке-мишени через Fс
- фрагмент молекулы иммуноглобулина,
взаимодействующий с Fc
- рецептором на мембране МФ
Опухолевая
клетка-мишень, экспрессирующая СТОА АТ
к СТОА МФ,
армированный АТ к СТОА
Специфическое
прикрепление МФ к клетке-мишени через
Fab - фрагмент
молекулы иммуноглобулина, взаимодействующий
со СТОА МФ Опухолевая
клетка-мишень Опухолевая
клетка-мишень
Киллерный
эффект МФ
в реализации противоопухолевой защиты
не сводится ни к их фагоцитарной
активности, ни к внеклеточному цитолизу,
обусловленному секретируемыми
продуктами. Более важную роль играют
механизмы, требующие прямого клеточного
контакта. Для достижения такого контакта
используются по крайней мере 3 механизма
(А.А. Ярилин,
1999):
А.
Неспецифический контакт, основанный
на взаимодействии адгезивных молекул
типа интегринов (LFA-1
— ICAM-1).














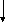



















































РОЛЬ МАКРОФАГОВ В ПРОТИВООПУХОЛЕВОЙ ЗАЩИТЕ
В. Взаимодействие Fc и CR-рецепторов МФ с опсонизирующими субстанциями на поверхности клетки-мишени (с антителами, компонентами комплемента). При этом МФ приобретает способность к антителозависимому цитолизу.
С. Специфическое взаимодействие с клетками-мишенями МФ, "армированных" путем связывания с их поверхностными Fc-рецепторами антител к клеткам-мишеням. В формировании "армированных" макрофагов важная роль принадлежит Т-клеткам и их продуктам, в частности, γ-интерферону.
Наибольшим киллерным эффектом обладают МФ, участвующие в реализации механизма С. Эффективность цитолиза в этом ряду клеточных взаимодействий с участием макрофагов последовательно повышается:
С > В > А
Природа киллерного эффекта МФ не раскрыта. Вероятно, как в и случае киллерных лимфоцитов, в основе киллерного действия МФ лежит сочетание различных механизмов (А.А. Ярилин, 1999):
индукция апоптоза;
внедрение в мембрану клетки-мишени цитолитических молекул, продуцируемых МФ;
продукция цитокинов с цитолитической активностью (ФНОα);
активные формы кислорода, галоидные производные и некоторые ферменты, секретируемые в межклеточную среду активированными макрофагами;
допускается возможность "инъекции" в клетку-мишень лизосом макрофагов.
Киллерной активностью обладают также гранулоциты. Если для эозинофилов это исключительно внеклеточный цитолиз, обусловленный секретируемыми продуктами, то для нейтрофилов, как и для макрофагов, киллерная активность связана с действием нескольких механизмов:
контактной индукцией апоптоза;
токсичностью секретируемых продуктов;
передачей в клетки-мишени токсического материала.
Естественные киллеры
(А.А. Ярилин, 1999; Р.М. Хаитов с соавт., 2000)
Для реализации активности NK не требуется их предварительного контакта с индуцирующим агентом и развития иммунного ответа. Они лишены антигенраспознающих рецепторов. В фило- и онтогенезе система естественных киллеров появляется раньше, чем факторы антигенспецифической иммунной защиты. Данная система ответственна за противоопухолевую и противовирусную резистентность организма.
В отличие от Т-лимфоцитов, NK-лимфоциты убивают клетки-мишени быстро (1-2 часа), используя упрощенный механизм без распознавания индивидуальных антигенов и подготовки в виде иммунного ответа.
Рецептор NK-клеток, предназначенный для распознавания клеток-мишеней представляет собой С-лектин, т.е. белок, распознающий углеводные остатки при участии ионов Са2+. Этот рецептор - NK R - Р1 (CD 161) распознает концевые остатки маннозы на молекулах мембранных гликопротеинов и гликолипидов. В норме такие остатки на большинстве клеток, с которыми контактируют зрелые лимфоциты и макрофаги, блокированы молекулами сиаловой кислоты. Это защищает их от фагоцитоза макрофагами (которые также имеют рецепторы, связывающие маннозу) и от лизиса NK.
Свободная манноза присутствует на поверхности юных и старых клеток, а также на пролиферирующих и трансформированных клетках.
Способность убивать клетки опухолевых линий у NK ограничена. Механизм этого ограничения связан с возможностью распознавания последними аутологичных молекул MHC при участии соответствующих рецепторов (KIR: NKB-1). В случае такого распознавания в NK-клетку передается сигнал, запрещающий развитие дальнейших событий, ведущих к цитолизу. В результате мишенями NK могут быть клетки, на поверхности которых присутствуют глюкоконъюгаты со свободными остатками маннозы и не содержатся молекулы МНС I класса (мера, предупреждающая повреждение собственных клеток организма).
При соблюдении этих условий происходит сближение NK-клеток с их мишенями, которое определяется не только связыванием рецепторов к глюкоконъюгатам, но и адгезивными взаимодействиями, в реализации которых принимают участие ганглиозиды, интегрины и другие молекулы.
После установления подобного контакта запускается программа апоптоза клетки-мишени с участием перфорин-зависимого механизма.
NK-клетки способны осуществлять рециклинг, т.е. отсоединяться от погибшей клетки и участвовать в повторном киллинге новых клеток-мишеней.