

Глава 3 / местные и общие реакции организма на повреждение
85
совокупности называемые антиоксидантами (табл. 10).
3.1.8. Стабильность липидного слоя мембран и явление электрического пробоя
Стабильность липидного слоя и электрический пробой мембраны. В отличие от белков и нуклеиновых кислот, которые в клетке собираются каждый по своему чертежу сложнейшими молекулярными роботами, липидный слой мембраны обладает способностью собираться сам по себе из молекул фосфолипидов и холестерина, если только они содержатся в водном растворе в достаточной концентрации. Это связано с особым свойством молекул липидов, входящих в состав мембран, которое принято называть ам-фифильностью, т. е. сродством одновременно к воде (гидрофильность) и к неводным средам, таким как растительное масло или жидкий парафин (гидрофобность). Молекула фосфолипида (основной липид клеточной и внутриклеточных мембран) имеет форму сплющенного цилиндра (рис. 13, А), один (меньший) конец которого хорошо растворяется в воде («полярная голова»), а другой - в воде не растворяется («жирный хвост») (рис. 13, Б). В водной среде такие молекулы самособираются в липидный бислой (рис. 13, В), который сам на себя замыкается, образуя везикулы - липосомы (рис. 13, Г).
Под влиянием тепловых движений молекул в липидном слое могут образоваться дефекты, которые приводят к образованию заполненных водой трещин и щелей (назовем их «порами»). Через такие дефекты могут проходить водорастворимые молекулы и ионы. Однако их появление крайне невыгодно с энергетической точки зрения, поскольку при этом граница раздела липид - вода сильно увеличивается, а это требует затраты работы на преодоление силы поверхностного натяжения. С ростом радиуса поры энергия системы растет пропорционально радиусу в соответствии с уравнением:
АЕ = nrlo,
где г - радиус поры; Z ■ толщина мембраны; а -энергия образования границы раздела площадью 1м2 (в системе СИ).
При наличии мембранного потенциала (т. е. разности потенциалов между водными фазами
86
по сторонам мембраны), который обозначается как фт, энергия образования поры снижается. Как показывает теория, в этом случае энергия системы изменяется с ростом поры по уравнению:
АЕ = nrlo ■ кг2 еоФ*т (s„ - ej/ 21,
где е0 - диэлектрическая постоянная, ец , ет - диэлектрическая проницаемость для воды и липидного слоя мембран соответственно; <р/( - мембранный потенциал.
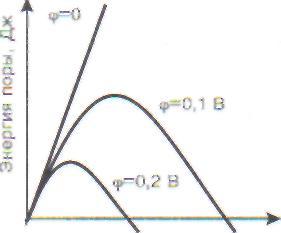
Изменение энергии поры с ростом ее радиуса при трех разных мембранных потенциалах показано на рис. 14, В. Видно, что с ростом радиуса энергия системы сначала растет, а затем начинает уменьшаться. Это означает, что после преодоления некоторого энергетического барьера рост поры будет происходить самопроизвольно, пока мембрана вообще не разрушится. Величина барьера снижается при увеличении мемб-

«Полярная голова»

Липидный бислой

«Жирный хвост»
г Липосома
Рис. 13. Самосборка фосфолипидов в бислой
Часть I. Общая нозология
рвнного потенциала. При небольших потенциалах, существующих в живой клетке (70 мВ на щдаоплазматической мембране и 175 мВ на внутренней мембране митохондрий), этого не проис-ходит, потому что барьер достаточно высок.
С ростом потенциала может наступить момент, когда в мембране начнут формироваться и расти ■оры и она будет разрушена. Такое явление ноет название электрического пробоя мембраны.

О
Образование водной поры
Поверхность поры равна 2кН
<
с
о I-
Радиус поры, мкм
0 40 80 120
_
Рис. 14. Электрический пробой мембран: А - появление в липидном бислое мембраны поры, заполненной водой; В - размер внутренней поверхности поры пропорционален ее радиусу; В - энергия мембраны с
порой в зависимости от ее радиуса. Величина потенциального барьера при росте поры уменьшается при увеличении потенциала на мембране; Г - возрастание тока в зависимости от потенциала пробоя
Величина потенциала, при котором начинается электрический пробой, называется потенциалом пробоя и обычно обозначается как U* или ф*. Величина потенциала пробоя, несколько различающаяся для мембран с разным составом белков и липидов, может служить количественной мерой электрической стабильности мембраны. Чем стабильнее мембрана, тем выше потенциал, который ее «пробивает» (т. е. ф*) .
Электрическая прочность различных мембранных структур. Явление электрического пробоя мембран изучалось многими авторами на искусственных мембранах и отдельных клетках. Мембраны обладают определенным сопротивлением R электрическому току I, которое при небольшой разности потенциалов ф между двумя сторонами мембраны является постоянной величиной. Иными словами, для мембраны соблюдается закон Ома:
I =Ф/Л.
Это означает, что зависимость между напряжением на мембране ф и током через мембрану I - линейная. Однако такая зависимость сохраняется при сравнительно небольших величинах ф: обычно не выше 200-300 мВ. При определенной разности потенциалов на мембране (потенциале пробоя ф*) происходит резкое возрастание тока (рис. 14, Г). При постоянном мембранном потенциале, если он превышает критическое значение, ток самопроизвольно нарастает во времени до полного разрушения мембраны.
На рис. 14 представлены результаты опыта на бислойных липидных мембранах. Аналогичные опыты были проведены на везикулярных мембранных структурах: фосфолипидных везикулах - липосомах, изолированных митохондриях и эритроцитах. В случае липосом и эритроцитов потенциал на мембране создавался за счет разности концентраций проникающих ионов по сторонам мембраны, в случае митохондрий - за счет энергии окисления субстратов. Измерение мембранного потенциала осуществлялось различными способами, например в случае митохондрий, - с помощью потенциалчувствительного флуоресцентного зонда. Явление пробоя мембран наблюдалось во всех случаях. В табл. 11 приведены величины потенциалов пробоя мембран всех этих объектов. Разумеется, потенциал пробоя во всех случаях выше потенциала, существующего на мембранах в живой клетке: иначе