

1.6 Метаболізм гліцеролу
Гліцерол, який вивільняється під час гідролізу триацил-гліцеролів або гліцерофосфоліпідів, захоплюється переважно печінкою і метаболізує за такими шляхами (рис.7):
окиснюється в аеробних (1) чи анаеробних умовах (2);
використовується як субстрат для синтезу глюкози в ГНГ(3);
повторно використовується для біосинтезу різних класів гліцеридів (4).

 Активація
Активація
Гліцеролкіназа
Г
 ліцерол
Гліцерол-3-Ф
Гл
ліцерол
Гліцерол-3-Ф
Гл
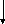

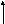
 АТФ
АДФ
НАД+
АТФ
АДФ
НАД+
(4) Гліцерол-3-Ф-ДГ (3)
НАДН2
 Синтез ТАГ, ФЛ
ДАФ
ГАФ
Синтез ТАГ, ФЛ
ДАФ
ГАФ
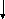
(2)

 Лактат ПВК
Лактат ПВК
(1)



 АТФ
Дихальний ланцюг ЦЛК
АТФ
Дихальний ланцюг ЦЛК
Рисунок 7 – Загальна схема шляхів метаболізму гліцеролу
Активація гліцеролу. Включенню гліцеролу до метаболіч-них шляхів передує його активація, тобто перетворення на
гліцерол-3-фосфат під дією ферменту гліцеролкінази:
Н2С – ОН Н2С – ОН
│ │
НС – ОН + АТФ НС – ОН + АДФ
 │Гліцеролкіназа
│
│Гліцеролкіназа
│
Н2С – ОН Н2С – ОРО3Н2
Гліцерол Гліцерол-3-Ф
Гліцерол-3-Ф здатен окиснюватися ферментом α-гліцеро-фосфатдегідрогеназою до діоксиацетонфосфату (ДАФ):
Н2С – ОН Н2С – ОН
│ НАД+ НАДН+Н+ │

 НС – ОН
С=О
НС – ОН
С=О
│ α-Гліцерофосфатдегідрогеназа │
Н2С – ОРО3Н2 Н2С – ОРО3Н2
Гліцерол-3-Ф Діоксиацетонфосфат
Діоксиацетонфосфат під дією тріозофосфатізомерази пере-творюється у гліцеральдегід-3-фосфат (ГАФ) - один із центральних метаболітів гліколізу.
Н2С – ОН О=С – Н
│ │

 С=О
НС – ОН
С=О
НС – ОН
│ Тріозофосфатізомераза │
Н2С – ОРО3Н2 Н2С – ОРО3Н2
Діоксиацетонфосфат Гліцеральдегід-3-фосфат
Подальше перетворення ГАФ іде по шляху ГНГ до Гл і глікогену, або по гліколітичному шляху до ПВК і далі по загальному шляху катаболізму до СО2 і Н2О (рис. 7).
Окиснення гліцеролу в анаеробних умовах призводить до виділення 2 молекул АТФ (як і в гліколізі), але враховуючи,
що 1 молекула АТФ використовується для активації гліце-ролу, енергетичний баланс становить - 1АТФ.
При повному аеробному окисненні гліцеролу до СО2 і Н2О енергетичний баланс становить 22 молекули АТФ.
Розрахунок:
а) 9 АТФ – утворюється за рахунок 3 НАДН2:
НАДН+Н+ відновлюється при окисненні гліцерол-3-фосфату до ДАФ;
НАДН+Н+ - з гліцеральдегід-3-фосфат-дегідроге-назної реакції;
НАДН+Н+ – з піруватдегідрогеназного комплексу.
б) 2 АТФ синтезуються за рахунок двох реакцій субстрат-ного фосфорилювання в гліколізі.
в) Окиснення 1 молекули ацетил-КоА до СО2 і Н2О в ЦЛК дає 12 АТФ.
г) 1 АТФ використовується на активацію гліцеролу у першій реакції.