

3 Швидкість синтезу жирних кислот контролюється енергетичним станом клітини (співвідношенням атф/адф).
3.1 Високі концентрації АТФ стимулюють синтез жирних кислот і, навпаки, переважання вмісту АДФ гальмує цей процес.
2.1.2 Елонгація жирних кислот. Утворення моно- і поліненасичених жирних кислот
Елонгація жирних кислот. Пальмітинова кислота (С16), яка утворюється у циклі Лінена, є попередником у синтезі довголанцюгових жирних кислот (С18, С20, С22, С24). Подовження ланцюга жирної кислоти відбувається за рахунок ензиматичної системи елонгації, яка має назву елонгаза жирних кислот і локалізується у цитозолі та мітохондріях клітини.
Утворення довголанцюгових жирних кислот відбувається шля-хом послідовного приєднання до ацильних радикалів двовугле-цевих фрагментів:
а) Мікросомальна система елонгації використовує малоніл-КоА як джерело двовуглецевих фрагментів і працює за механізмом, подібним до синтази ВЖК:
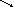 (малоніл-КоА,
НАДФ) С2
Елонгаза
(малоніл-КоА,
НАДФ) С2
Елонгаза
П



 альмітоїл-КоА
(С16)
Стеароїл-КоА
(С18)
альмітоїл-КоА
(С16)
Стеароїл-КоА
(С18)
Субстратами мікросомальної елонгази є насичені жирні кислоти з С10 та більшою кількістю атомів вуглецю.
б) Мітохондріальна система елонгації використовує ацетил-КоА як донора двовуглецевих фрагментів і подовжує С12-С16 жирні кислоти:
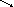 ацетил-КоА
(С2)
Елонгаза
ацетил-КоА
(С2)
Елонгаза
 ацил-КоА
(Сn)
ацил-КоА (Сn+2)
+ КоА-SH
ацил-КоА
(Сn)
ацил-КоА (Сn+2)
+ КоА-SH
Утворення моно- і поліненасичених жирних кислот
Організм людини має обмежені можливості щодо перетворення насичених жирних кислот у ненасичені. Мононенасичені жирні кислоти пальмітоолеїнова (С16:1) та олеїнова (С18:1) можуть утворюватися в організмі людини з відповідних насичених жирних кислот – пальмітинової (С16) та стеаринової (С18). Ці перетворення відбуваються у мікросомах печінки і жирової тканини за участю системи десатурації жирних кислот (ацил-КоА-оксигенази), яка за механізмом дії є цитохром b5-вмісною монооксигеназою та утворює подвійний зв'язок між 9-м і 10-м атомами вуглецю (Δ9).
Процеси десатурації (утворення подвійних зв'язків) та елонгації (подовження ланцюгів) можуть сполучатися і повторюватися, що дає можливість синтезувати різноманітні мононенасичені жирні кислоти з довгими ланцюгами (рис. 11).
СН3-(СН2)7-СН=СН-(СН2)7-СО~S-КоА
Олеїл-КоА (С18:1)
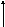
 2Н2О
+ НАДФ+
2Н2О
+ НАДФ+
Десатураза
О2+ НАДФН·Н+
СН3-(СН2)16-СО~S-КоА
Стеароїл-КоА (С18:0)

 (малоніл-КоА,
НАДФ·Н+)
(С2)
Елонгаза
(малоніл-КоА,
НАДФ·Н+)
(С2)
Елонгаза

СН3-(СН2)14-СО~S-КоА
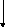 Пальмітоїл-КоА
(С16:0)
Пальмітоїл-КоА
(С16:0)
 О2
+ НАДФН·Н+
О2
+ НАДФН·Н+
Десатураза
2Н2О+ НАДФ+
СН3-(СН2)5-СН=СН-(СН2)7-СО~S-КоА
Пальмітоолеїл-КоА(С16:1)
Рисунок 11 – Шляхи перетворення пальмітинової кислоти у реакціях
десатурації та елонгації
На відміну від рослин, у клітинах людини і тварин відсутні десатурази, які утворюють подвійні зв'язки після 9-го атому вуглецю. З цієї причини в організмі людини не можуть синтезуватися поліненасичені жирні кислоти – лінолева С18:2(Δ9,12), ліноленова С18:3(Δ9,12,15). Ці кислоти відносяться до категорії незамінних (есенційних) і повинні постійно надходити до організму з їжею. Незамінні жирні кислоти під дією систем десатурації та елонгації, що містяться у ендоплазматичному ретикулюмі гепатоцитів, трансформуються у такі поліне-насичені жирні кислоти, як γ-ліноленова С18:3(Δ6,9,12), ейкоза-трієнова С20:3(Δ8,11,14), арахідонова С20:4(Δ5,8,11,14), докозан-гексенова С22:6. До незамінних жирних кислот також відносять арахідонову кислоту, яка утворюється з лінолевої (рис.12).
 Лінолеїл-КоА
С18:2(Δ9,12)
Лінолеїл-КоА
С18:2(Δ9,12)
 О2
+ НАДФН·Н+
О2
+ НАДФН·Н+
Δ6-Десатураза
2Н2О+ НАДФ+
 γ-Ліноленіл-КоА
С18:3(Δ6,9,12)
γ-Ліноленіл-КоА
С18:3(Δ6,9,12)
 С2
С2
(малоніл-КоА, НАДФ·Н+) Мікросомальна
елонгаза
Дігомо- γ-Ліноленіл-КоА С20:3(Δ8,11,14)
О2
+ НАДФН·Н+
 Δ5-Десатураза
Δ5-Десатураза
Н2О+ НАДФ+
Арахідоніл-КоА С20:4(Δ5,8,11,14)
Рисунок 12 – Схема перетворення лінолевої кислоти в арахідонову
За умов надходження до організму достатньої кількості ліно-левої кислоти, потреби людини у арахідоновій кислоті повністю задовольняються. Арахідонова кислота є попередником у синтезі біологічно активних речовин - ейкозаноідів.
При голодуванні та нестачі інсуліну процеси елонгації та десатурації суттєво зменшуються.
Відсутність або нестача незамінних жирних кислот в їжі протягом довгого часу, яка може спостерігатися у немовлят, що знаходяться на штучному вигодуванні, або у хворих, життєдіяльність яких підтримується лише за рахунок парентерального харчування, призводить до відставання у рості, розвитку дерматиту. Для запобігання таких ускладнень кількість незамінних жирних кислот повинна становити не менше ніж 1-2% від загальної потреби організму у калоріях.