

1.3.2 Молекулярні механізми регуляції ліполізу
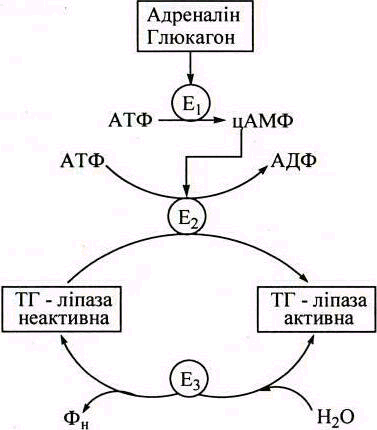
Загальна швидкість ліполізу контролюється активністю ТАГ-ліпази - регуляторного ферменту, який каталізує найбільш повільну реакцію ліполізу. Регуляція активності ензиму відбувається через каскадний механізм за допомогою гормонів: адреналіну, глюкагону, інсуліну, соматотропіну (рис.2).
Активність двох останніх ензимів значно вища і залежить від концентрації відповідних субстратів реакцій.


(
 Е1)
– аденілатциклаза;
(
+)
Е1)
– аденілатциклаза;
(
+)
(
 Е2)
- протеїн-кіназа;
ТАГ
ДАГ+ ЖК
Е2)
- протеїн-кіназа;
ТАГ
ДАГ+ ЖК
(ЕЗ) - протеїнфосфатаза. ДАГ- ліпаза
МАГ + ЖК 
МАГ- ліпаза
Гліцерол + ЖК
Рисунок 2 – Каскадна регуляція активності ТАГ-ліпази адипоцитів
Молекулярною основою регуляції активності ТАГ-ліпази адипоцитів є її ковалентна модифікація (обернене фосфорилю-вання-дефосфорилювання). Каталітично активною є фосфори-льована форма ТАГ-ліпази, дефосфорильована - неактивна.
1.3.3 Нейрогуморальна регуляція ліполізу
Катехоламіни (адреналін, норадреналін), глюкагон - активують ліполіз за рахунок стимуляції цАМФ-залежної каскадної системи і активації ТАГ-ліпази адипоцитів. Ліполітична дія катехоламінів реалізується при фізичному напруженні, стресі, зниженні температури навколишнього середовища. Дія глюкагону проявляється при зменшенні надходження глюкози через шлунково-кишковий тракт або посиленні використання її тканинами. За рахунок дії катехоламінів та глюкагону відбувається швидка активація ліполізу в жировій тканині та мобілізація НЕЖК, що забезпечує підвищені енергетичні потреби організму за умов стресу або голодування.
Інсулін інгібує ліполіз і вивільнення ЖК із адипоцитів за рахунок двох біохімічних механізмів:
а) зменшення концентрації цАМФ через активацію фосфодіесте-
рази цАМФ;
б) підвищення проникності мембран адипоцитів до глюкози, що
приводить до активації гліколізу і ліпогенезу з продуктів
метаболізму глюкози.
Соматотропін - активує ліполіз у жировій тканині при голодуванні за рахунок підвищення синтезу відповідних ферментних білків. Метаболічні ефекти соматотропіну розвиваються повільно.
Ліполіз в інших тканинах (м'язах, печінці) регулюється за подібними нейрогуморальними механізмами.
1.4 Окиснення жирних кислот: послідовність реакцій, енергетика процесу
У результаті ліполізу ТАГ, з жирової тканини вивільняються жирні кислоти і гліцерол. Гліцерол метаболізує у печінці, жирні кислоти окиснюються в різних тканинах з генерацією значної кількості АТФ.
1.4.1 Послідовність реакцій окиснення жирних кислот
Окиснення жирних кислот відбувається у матриксі мітохондрій у циклічному процесі, який має назву b-окиснення жирних кислот (окиснюється b-атом вуглецю жирної кислоти). Відбувається послідовне відщеплення від молекули насиченої жирної кислоти з парною кількістю атомів вуглецю двохвуглецевих фрагментів Ацетил – КоА (рис. 3).
(1) активація (2) транспорт (3) b-окиснення
Ж




 К
Ацил-КоА
Ацил-КоА
Ацетил-КоА
К
Ацил-КоА
Ацил-КоА
Ацетил-КоА


 ФАДН2
і НАДН2
(4)
ЦЛК
ФАДН2
і НАДН2
(4)
ЦЛК

 Дихальний
ланцюг
Дихальний
ланцюг