

Блохин Г.И., Александров В.А. Зоология
.pdfРис. 27. Инфузории:
4 I ус|х-лька; б — трубач; в — брюхоресничная инфузория, ползающая с помощью утолщенных щетинок; г — хищные инфузории колепсы, напавшие на другую инфузорию; I нгрздний конец; 2 — пищеварительные вакуоли; 3 — микронуклеус; 4 — реснички; I к «с точная глотка; 6 — удаленная через порошицу непереваренная пища; 7— сокра-
• ик'/ила» вакуоль; 8—макронуклеус; 9—трихоцисты; 10—приводящие каналы сокра1ИИ И1.НОЙ вакуоли
МI югие инфузории обладают сложной системой пищеварения. Рас- моможепный в углублении тела (перистоме) рот, или цитостом, окру- •гн длинными ресничками, с помощью которых пищевые частички и 1Я1 пииются в него. Часто рот ведет в глотку, погруженную в эндоплазму И )i 1доплазме пищевые частицы окружаются пузырьками, содержа- 11|ими пищеварительные ферменты, в результате образуются пищева- рительные вакуоли. В пищеварительных вакуолях создается кислая аре да. На последующих этапах переваривания пищи среда становится ни*ночной, что аналогично процессам пищеварения у высших живот- ных Непереваренные остатки пищи выбрасываются из тела инфузории н определенном месте через порошицу. Есть виды хищных инфу- *»|>И й, вооруженных ротовым хоботком, с помощью которого они промныиают покровы своей жертвы.
Большинство инфузорий питаются бактериями, немногие поедают одноклеточные водоросли, и среди них встречаются даже монофито- ||ш1 и. Хищные инфузории порой охотятся на жертв, которые по разме- рим больше хищниц; это, например, инфузория-туфелька. Жертву они поражают хоботком и высасывают ее содержимое. Свободноживущие инфузории являются важным звеном в пищевых цепях экосистем.
51
У пресноводных инфузорий имеются сократительные вакуоли, выполняющие функции осморегуляции и выделения. Иногда сократительные вакуоли усложнены несколькими приводящими каналами. В этих каналах накапливается избыток жидкости, которая затем выбрасывается в основной резервуар; из него жидкость выталкивается за пределы тела одноклеточного.
Инфузории размножаются бесполым путем, делясь надвое, но в поперечном направлении. При этом ядро делится с помощью митоза. Половой процесс в виде конъюгации не приводит к увеличению числа особей. При конъюгации две размножающиеся особи соединяются попарно. Перед соединением в каждой особи разрушаются макронуклеусы, а микронуклеусы делятся мейотически, образуя четыре гаплоидных ядра. Из них три также разрушаются, а четвертое митотически делится на два ядра. Одно из этих ядер остается в клетке, а второе мигрирует в другую особь. После такого обмена ядрами оставшееся в клетке стационарное ядро сливается с ядром-мигрантом. В результате образуется диплоидное ядро. Затем конъюгирующие особи расходятся.
У каждой особи после расхождения из диплоидного ядра формируется макронуклеус и микронуклеус. Таким образом, в результате конъюгации в каждой особи изменяется генотип.
При классификации инфузорий в качестве диагностических признаков используют особенности строения ротового аппарата или структуру ресничного аппарата. Последний подход преобладает. Инфузорий делят на два класса: класс Ресничные инфузории (Ciliata) и класс Сосущие инфузории (Suctoria).
Класс Ресничные инфузории наиболее многочисленный. Представители этого класса покрыты ресничками на протяжении всех стадий жизненного цикла. Среди представителей подкласса Равноресничные инфузории (Holotricha), характеризующихся равномерным расположением на теле ресничек равной длины, много свободноживущих (например, инфузория-туфелька, Paramecium caudatum), хищных, питающихся своими собратьями, и паразитических форм. Среди последних следует отметить инфузорию балантидий (Balantidium coli), которая встречается в кишечнике свиней и человека. Эта инфузория питается в основном содержимым кишечника, но может разрушать слизистую кишечника, вызывая заболевание — балантидиоз. Заражение происходит при потреблении загрязненного цистами балантидия пищи и воды.
В природных водоемах и в прудовых хозяйствах, занимающихся разведением рыб, большой вред наносят паразитические инфузории. Например, равноресничная инфузории Ichthyophthirius внедряется в кожу рыб и начинает питаться клетками хозяина. В результате на теле рыбы образуются многочисленные язвочки. Заболевание может привести к гибели рыб, особенно молоди карпа. На жабрах и коже часто паразитируют инфузории из рода Trichodina, причиняя молоди рыб существенный вред.
У большинства кругоресничных инфузорий (подкласс Кругоресничные инфузории — Peritricha) реснички располагаются левоспи-
52

Рис. 28. Сидячие инфузории сувойки:
l,J — деление сувойки; 3 — плавающая, отделив-
|||»игн от материнского организма клетка-бро- ДКМхл; 4— половой процесс
риш.но только вокруг предротовой во- ронки. Многие формы ведут прикреп- ленный образ жизни. Так, Сувойки (Vortfcella, отр. Peritrichida) имеют винный сократимый стебелек, с по- мощью которого они прикрепляются к суОс грату. Среди них есть и колониаль- ные формы (рис. 28).
У спиральноресничных инфузорий (подкласс Спиральноресничные инфу-
зории — Spirotricha) полоса ресничек, ведущих ко рту, закручена впра- ии Среди этих одноклеточных особое место принадлежит инфузоримм, живущим в рубце жвачных животных (отр. Entodiniomorpha). Их Т»ло одето в прочный панцирь с многочисленными отростками (рис. 29), поэтому их называют панцирными инфузориями. Питаются они бактериями рубца и способствуют расщеплению клетчатки кор- ми, взаимодействуя сложным образом с целлюлозорасщепляющими Оик гериями. Эти полезные симбионты не только участвуют в перева- риипнии пищи, но и сами служат источником питания для жвачных «и потных.
11ростейшие из класса Сосущие инфузории (Suctoria) лишены ресни- чек I in большей части жизненного цикла, и лишь на ранних этапах раз- пи гии дочерняя клетка — бродяжка имеет реснички. У них нет рта и околоротовой воронки. Эти инфузо- рии с шаровидным телом, на котором иидиально расположены щупальца {рис. 30). Щупальца служат для при- крепления к субстрату и, кроме того, нплнются ловчим аппаратом. Поймав мелких инфузорий с помощью липкого секрета, суктории как бы перекачива- ют содержимое жертвы в свое тело.
При бесполом размножении от ма- (ерииской клетки суктории отпочкопымнется дочерняя особь — бродяжка,
Ги< 24. Инфузория из желудка жвачного млекопитающего:
I перепонки, загоняющие пищу в рот; 2— клеfO'llinit глотка; 3 — реснички; 4 — сократительные никуоли
53
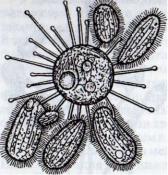
Рис. 30. Сосущая инфузория Sphaerophrya, высасывающая щупальцами содержимое нескольких ресничных инфузорий
покрытая ресничками. В последующем она теряет ресничный аппарат и превращается в сукторию с щупальцами.
Следует отметить важную роль некоторых мелких видов инфузорий, жгутиконосцев и амеб в жизни почвы. Бактерии, в том числе азотфиксирующие, служат основной пшцей для простейших. Однако они не только поедают бактерий, но и способствуют их раз-
множению, выделяя в почву вещества, стимулирующие этот процесс. Простейшие способствуют повышению плодородия почвы, активно участвуют в процессах биологической очистки водоемов.
Имеется опыт искусственного разведения паразитических простейших для борьбы с вредными насекомыми.
ФИЛОГЕНИЯ И ЭКОЛОГИЧЕСКАЯ РАДИАЦИЯ ПРОСТЕЙШИХ
Мир живых существ делят на два надцарства: Безъядерные, или Прокариоты (Procaryota), и Ядерные, или Эукариоты (Eucaryota).
Клетки прокариот в отличие от клеток эукариот не имеют оформленного ядра. Эукариот обычно делят на три царства: Растения (Vegetabilia, или Plantae), Грибы (Mycetalia, или Fungi) и Животные (Animalia, или Zoa). Большинство растительных организмов — автотрофы, самостоятельно синтезирующие органические вещества в процессе фотосинтеза. Грибы, хотя и относятся к гетеротрофным организмам, но питаются растворенными органическими веществами. Животные являются гетеротрофами, существующими за счет потребления других организмов или их остатков. Но различия по типам питания между этими царствами относительны, так как имеется множество переходных форм, особенно среди низших представителей.
Простейших (Protozoa) относят к примитивным одноклеточным эукариотам. Признано, что эукариоты берут начало от прокариот. Косвенным свидетельством их единства служит сходство процессов синтеза белка в клетке. Безъядерные организмы были одними из первых на нашей планете, часть из них способна существовать даже в бескислородной среде.
Среди эукариотических животных клеточный уровень организации рассматривают как более примитивный. Это позволяет полагать, что простейшие на Земле появились первыми и послужили началом более слож-
54
UttR форм — многоклеточным животным. В настоящее время простей- существуют во взаимодействии с более сложными по организа-
ЦИи многоклеточными организмами. Этому способствует то, что Бритсйшие в процессе эволюции отлично приспособились к разноЛЦМЫНЫм условиям жизни на нашей планете,
и Полагают, что эукариоты произошли от прокариот путем постеМиного возникновения органелл из мембраны клетки прокариот. В ниолюции эукариот, по-видимому, большую роль сыграл симбиоз
&ЯЛИЧНЫХ прокариот. Из известных семи типов простейших четыре |ЙЛй представляют исключительно паразитические группы, которые
&М1Икли значительно позднее, после появления их хозяев — высших РМотклеточных. Поэтому при выяснении наиболее примитивной
Шумны среди простейших все внимание уделяется таким типам, как
Свркомастигофоры и Инфузории.
I ИI |фузорий обычно относят к самым высокоорганизованным одно- Ийгтчным животным: есть представители с чертами многоклеточно- (ТИ. Таким образом, только саркомастигофоры несут первичные при- емки, имеющие сходство с предками всех простейших. Но саркомастиЩфоры чрезвычайно неоднородны, что подтверждается особенностью (ргпнизации подтипов Саркодовые и Жгутиконосцы.
Кольшинство исследователей придерживаются мнения Пашера 1|914)о том, что жгутиковых следует относить к более древней группе.
' них много общего с одноклеточными растениями, разнообразнее тиNM иитания, аих органеллы движения имеются даже у части прокариот. Можно предположить, что разнообразные способы питания у жгути- Иопых могли стать основой для последующего их разделения на авто- )фоп и гетеротрофов. Следует отметить, что жгутики есть у гамет ilozoa и Metazoa. Упрощенность (отсутствие жгутиков и пелликулы) •ргннизации саркодовых можно отнести к вторичным явлениям в свя- 1И с переходом к особому типу питания — фагоцитозу Наличие же жгу- ТИКом у гамет части саркодовых может лишь свидетельствовать об их Происхождении от жгутиконосцев. Кроме того, в последнее время было Показано, что у некоторых амеб с ростом происходит редукция жгутиNOH у взрослых форм. Описано немало форм саркодовых, обладающих
Иовидоподиями и жгутиками одновременно.
Таким образом, можно предположить, что предками современных Иго! о/.оа были древние Саркомастигофоры с разными способами пита- Мим и имеющие примитивные жгутики.
бесспорно, что Apicomplexa и Ciliophora имеют родство со жгу- тиконосцами. Споровики могли упроститься в связи с переходом к Паразитическому образу жизни, но одновременно с этим у них усложнился жизненный цикл, который включает стадию гамет, снаб-
женных жгутиками. Myxozoa и Microspora могли произойти от древ- них саркодовых, поскольку их развитие начинается с амебоидного иродыша, а гаметы со жгутиками отсутствуют. Возможны вариан- ты пнтономности их эволюционного развития. На основе изложенного выше материала представляется возможной филогенетическая
55
|
|
|
|
|
схема Protozoa, представленная ни |
|||
|
|
я |
|
о. |
рис. 31. |
|
|
|
|
"8 |
я |
а. |
Е |
На |
базе современных |
пред |
|
|
с |
о |
||||||
о |
|
я |
о |
и |
ставлений можно обозначить ос |
|||
X |
10 |
|
а. |
|||||
|
а. |
|
< |
новные |
|
пути экологической |
||
|
W |
О |
|
|
|
|||
|
|
|
|
|
радиации |
одноклеточных |
ж и |
|
|
|
|
|
|
вотных (рис. 32). Центральном |
|||
|
|
|
|
|
группой могли быть многочис- |
|||
|
|
|
|
|
ленные и разнообразные предста |
|||
|
|
|
|
|
вители |
Саркомастигофор, |
но с |
|
|
|
|
|
|
преобладанием жгутиковых форм |
|||
|
|
|
|
|
и господством водных форм. В по- |
|||
|
|
|
|
|
следующем саркодовые утратили |
|||
|
|
|
|
|
жгутики и осуществили переход к |
|||
|
|
|
|
|
ползающему образу жизни и пита |
|||
|
|
|
|
|
нию путем фагоцитоза и пиноцито- |
|||
|
Примитивные |
|
|
за. Бентосный образ жизни способ- |
||||
|
Sarcomastigophora |
|
|
ствовал образованию защитных ра- |
||||
Рис. 31. Филогения Protozoa |
|
ковин разнообразной конструкции, |
||||||
|
|
|
|
|
таких как |
раковины корненожек, |
||
фораминифер. У части саркодовых наружный скелет усложнился и стал более легким, в результате чего они смогли перейти к планктонному образу жизни (радиолярии, солнечники).
В связи с прогрессом клеточного строения возникают крупные по лиэнергидные формы — инфузории. Они активно передвигаются, ведут разнообразный образ жизни. Среди них плавающие, ползающие, сидячие, скважники. Позже возникают симбиотические и затем паразитические формы инфузорий. Последние и поныне продолжают свое развитие, усложняя и совершенствуя отдельные этапы своего жизненного цикла.
Многим простейшим, в основном обитающим в пресных водоемах и паразитирующим в других организмах, свойственно образование цист и спор при наступлении неблагоприятных условий (высыхание и вымерзание водоемов). У морских представителей инцистирование является лишь исключением. В виде цист и спор простейшие могут переноситься ветром, птицами и другими животными на большие расстояния, благодаря чему пресноводные простейшие встречаются на земной поверхности повсюду, где имеются условия для их существования.
Пресноводные простейшие заселяют все виды водоемов, даже самые мелкие лужи или скопления воды, образующиеся в пазухах листьев растений. Множество их и в болотах, где в основном встречаются раковинные амебы. По составу фауны простейших относительно точно можно определить степень загрязнения водоема, так как в зависимости от этой степени водоемы заселяют разные представители простейших; их состав меняется по мере возрастания или снижения степени загрязнения воды. Простейшие живут даже в горячих источниках с темпера-
56
ш
Эндозообионты
амебоидные
Эндофитобионты
|
|
Эндозообионты |
|
|
|
|
жгутиковые, |
|
|
|
|
ресничные |
|
|
|
Бентос |
Эндозообионты |
|
|
|
|
|
|
|
|
|
Планктон |
|
|
.•лиобионть |
|
|
|
|
|м^овин"нь1е |
|
. ' . \.'/Интргаёен.тр(5"иййты |
||
|
|
|||
|
|
; Э.Лк/O^viQVITbl'•I .•;'.';•; ; :'. •'•'•'• '' |
||
|
|
.'•".'•.:си'дячйё ' . .'•.'• |
. ' . ' • |
|
|
|
;;:•'.• '.; • •Зли'о'ёионты' .. •'/'.'•''.'.'••' •'." |
:.'.•'•• •'.'.'• |
|
|
|
•'. /• .'эмфбйидиьье. '..'•• - .'. .'•••. J |
; '.'.• |
•'•'. '••.'.' • |
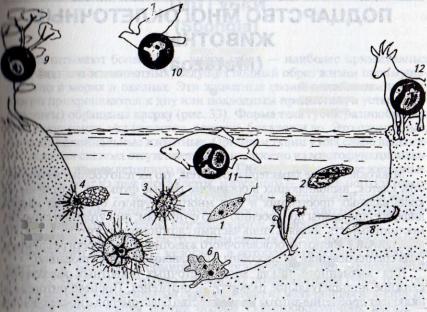
Рис. 32. Экологическая радиация простейших:
I *i ушконосец; 2 — инфузория; 3 — радиолярия; 4 — раковинная амеба; 5— форамиМмфпш; 6— амеба; 7—сувойка; 8— инфузория; 9 — лептомонас; 10— кишечная амеба; II, I/ различные паразитические простейшие
I v|niii поды до +50 "С, находят их и в водоемах с высоким уровнем заемкости (до 25 %).
Наиболее разнообразен состав простейших в морях и океанах, где они ни сияют в огромных количествах всю толщу воды. Вместе с одноклеточ- ми водорослями простейшие служат кормом для других животных, в HIM числе для рыб и китообразных. Раковинные саркодовые, отмирая, опускаются на дно, образуя мощнейшие донные отложения известняков.
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ ЖИВОТНЫЕ
(Metazoa)
Многоклеточные характеризуются тем, что их тело состоит из множества клеток, выполняющих в организме разные функции. В отличие от колониальных простейших в теле многоклеточного организма клетки дифференцированы по строению и по функциям; они утратили свою са мостоятельность, являясь лишь частями сложного единого организма. По этой причине клетки многоклеточного животного, приобретя различною рода специализацию, не способны к независимому существованию.
Для жизненного цикла многоклеточных животных характерно сложное индивидуальное развитие — онтогенез. Процесс онтогенеза включает дробление зиготы на множество бластомеров с последующей дифференцировкой их на зародышевые листки и зачатки органов, развитие, рост и образование взрослого организма. Увеличение размером тела многоклеточных по отношению к их поверхности способствовало усложнению процессов обмена веществ, что, в свою очередь, обеспечило многоклеточным животным устойчивость жизненных процессов и способствовало продлению их жизни.
Большинство ученых считают, что многоклеточные произошли от Protozoa. В пределах Protozoa прослеживается тенденция перехода к многоклеточности. В отдельных случаях у Protozoa наблюдается даже многоклеточность отдельных фаз развития (Миксоспоридии). Структурные компоненты клеток Protozoa идентичны таковым клеток Metazoa. Важное значение в решении вопроса о происхождении многоклеточных многие исследователи приписывают колониальным простейшим, например Volvox, у которого имеются клетки двух типов — соматические и половые.
Еще в 1874 г Э. Геккель утверждал, что предком многоклеточных была шаровидная колония какого-то простейшего, и что в процессе эволюции (филогенеза) за счет впячивания одной половины шара могла возникнуть первичная кишечная полость и первичный рот. Такой уже двухслойный организм плавал с помощью жгутиков, размножался половым путем и впоследствии стал предком многоклеточных.
Существует множество других теорий, но большая часть их сходится в одном: отдаленными предками многоклеточных животных были колониальные простейшие организмы.
58
ТИП ГУБКИ (Spongia)
Ншчитмпают более 5 тыс. видов губок — наиболее примитивных Milt мок неточных животных, ведущих водный образ жизни преимущенпенно и морях и океанах. Эти животные своим основанием неподнманп прикрепляются к дну или подводным предметам, а устьем (отмер* I нем) обращены кверху (рис. 33). Форма тела губок разнообразна: нмн могут иметь вид наростов, веточек, бокалов и др. Это одиночные, Но чище колониальные животные. Органы и ткани у них слабо выражены нгрпнли система отсутствует. Тело построено из многофункциональ-
ны* •пгтк и межклеточного вещества. Губки |
двухслойные животные. |
И* нчю состоит из двух слоев клеток: наружного |
дермального (эктодер- |
мн) и ину|рсннего — гастрального (энтодерма). Гастральный слой выстиНш1 инугреннюю (парагастральную) полость. Он состоит из так называе- мы» in ми» п шчковых клеток, или хоаноцитов, имеющих жгутики.
М г жду дермальным и гастральным слоями клеток имеется слой бес- I • |• у»I vpnoro вещества мезоглеи, в котором разбросаны отдельные ММ MI В мезоглее формируется органический или минеральный (( Й< (> I. Si()2) скелет; только у немногих представителей губок тело осHMMt н Mm ким. Минеральный скелет состоит из мельчайших игл-спикул, hoHipiae <|юрмируются внутри особых скелетообразующих клеток —
МроАлнстов, расположенных в мезоглее. Роговой (спонгиновый) ске- им но химическому составу близок к шелку.
I loiiepxnocTb тела губок пронизана множеством пор, через которые Миш поступает в систему каналов и камер. Движение воды обеспечиваем * I ушки хоаноцитов. Из каналов и камер вода попадает в центральную иирагастральную полость, откуда выводится наружу через устье.
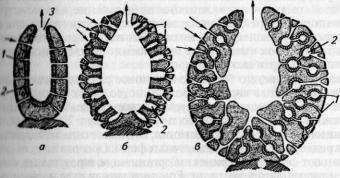
I'm II Тины строения губок с различной сложностью системы каналов и расположением
жгутиковых камер:
Л ш и ш . б сикон; в — лейкон; 1 — поры; 2 — жгутиковые камеры; 3 — устье(стрелммн у» и мно направление тока воды в теле губки)
59
Специальные клетки пороциты способны к сокращению и могут открывать и закрывать поры.
Среди клеток, находящихся в мезоглее, выделяются подвижные клетки амебоциты. По своим функциям они универсальны: выпол няют транспортную функцию, выделяют вещество мезоглеи, из них образуются половые клетки и клетки всех других типов, они участвуют м бесполом размножении, дают начало клеткам, образующим скелет жи вотного. Они переносят пищевые частицы от хоаноцитов к другим клеткам, удаляют экскреты, а в период размножения переносят спермин в мезоглее к яйцеклеткам. Такие разнообразные функции амебоцитов характеризуют губок как животных, находящихся на эволюционной лестнице ниже прочих многоклеточных.
Наиболее простой тип строения губок называют аскон (рис. 33), но он в основном характерен для одиночных форм и для молодых колониальных особей. Усложнение в период индивидуального развития приводит к возникновению более сложной формы — типу сикон. Дальнейшее усложнение строения тела губок (утолщается мезоглея, образуются карманы и камеры, покрытые слоем воротничковых клеток — хоаноцитов) ведет к самому сложному типу — лейкон. Таким образом, у губок типа лейкон и частично сикон парагастральная полость (в отличие от типа аскон) оказывается выстланной клетками эктодермы. Ими же выстланы приводящие и отводящие (у лейкон) каналы, которые являются впячиваниями эктодермы. Энтодермой выстланы лишь жгутиковые камеры, число которых у губок лейконового типа огромно — до нескольких миллионов.
Движение воды по каналам тела губок обеспечивает их организм кислородом и способствует удалению из тела продуктов обмена. С водой в тело губок попадают пищевые частицы (мелкие водные животные и растительные организмы, гниющие остатки), которые захватываются псевдоподиями хоаноцитов и перевариваются в их цитоплазме. Часть захваченной пищи передается амебоцитам, и те ее переваривают и транспортируют питательные вещества в мезоглею. Таким образом, губкам свойственно внутриклеточное пищеварение, в парагастральной полости пища не переваривается; эта полость служит лишь для сбора и эвакуации поступившей в организм губки воды. Существенное значение в питании губок имеет также поглощение растворенных органических веществ осмотическим путем.
Губкам свойственно бесполое и половое размножение. Среди них есть раздельнополые виды, но большинство губок — гермафродиты. Бесполое размножение осуществляется наружным почкованием или образованием особых внутренних почек — геммул. Если бесполое размножение носит характер почкования, то полное отделение почки происходит редко, в основном у одиночных форм. Обычно же дочерние особи сохраняют связь с материнским организмом, в результате чего образуются и разрастаются колонии. Границы между отдельными особями могут исчезать, и тогда вся колония сливается в общую массу (рис. 34).
Геммулы в виде групп клеток, окруженных оболочками и содержащих запас питательных веществ, образуются в мезоглее. Эти образова-
60