

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfÐèñ. 13.13. Избирательное включение пейсмекера микроаппликациями АХ в разные локусы хемочувствительной мембраны изолированного нейрона.а — отсутствие ответа при действии АХ в локус 1; б — кратковременное включение пейсмекерной активности при действии нейромедиатора в локус 2; в — активация пейсмекера и длительное сохранение его после нанесения АХ в локус 3. Калибровка: 10 мВ, 1 с.
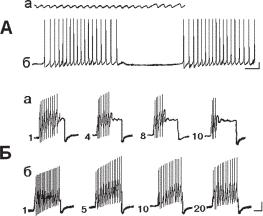
Ðèñ. 13.14. Взаимодействие фонового пейсмекера с синаптическими потенциалами. а — фоновая активность нейрона, зарегистрированного в полуинтактном препарате моллюска Helix lucorum; б — ответ нейрона на тактильное раздражение, примененное в область ноги моллюска. Калибровка: 10 мВ, 1 с.
Синаптическая регуляция пейсмекерной активности. Для выяснения природы медиаторного взаимодействия между интернейроном висцерального ганглия, инициирующим пейсмекерную активность в нейроне ППа1, и самим ППа1 нейроном у улитки Helix lucorum Кононенко и Осипенко (1987) были проведены специальные опыты. Найдено, что стимуляция интернейрона приводила к инициации берстовой активности в нейроне ППа1. Замена экстраклеточного Ca2+ на Mg2+, так же, как и добавление в физраствор во внешнем растворе, обратимо тормозит действие интернейрональной стимуляции на нейрон ППа1. Увеличение концентрации ионов внеклеточного Са2+ до 70 мМ не предотвращает этого действия. Внутриклеточный цезий Cs+ и инъекция тетраэтиламмония (ТЭА) в интернейрон, увеличивающие длительность потенциалов действия, также приводили к увеличению эффективности связи между нейронами. Эти результаты позволяют сде-
350
лать вывод о том, что связи между неронами моносинаптические и пептидергические.
Каковы постсинаптические механизмы связи между интернейроном и нейронами, имеющими пейсмекерную активность? В опытах Кононенко и Осипенко (1987) исследовалось взаимодействие между интернейроном, запускающим берстовую активность в идентифицированных клетках ППа1 и В7. В условиях фиксации потенциала регистрировался медленный входящий ток после стимуляции интернейрона. Снижение температуры приводило к снижению скорости развития и снижения этого тока с температурным коэффициентом около 10. Снижение концентрации ионов Na+ приводит к падению амплитуды. Был сделан вывод о том, что стимуляция интернейрона активирует по крайней мере два типа ионных каналов в соме берстовых нейронов. Предполагается,что этот процесс опосредован цепью биохимических цитоп-
лазматических реакций.
Модуляция пейсмекерной активности и синаптические связи. В опытах Кононенко и Сторожука (1992) на нейронах моллюска Helix pomatia в условиях фиксации напряжения на мембране изучали действие внешней аппликации дофамина на связи между пептидерги- ческим интернейроном, инициирующим берстовую пейсмекерную активность в неактивной клетке ППа1, и самим берстовым нейроном. Внешняя аппликаци дофамина по типу доза-эффект продуцирует обратимое торможение медленного входящего тока в ППа1 нейроне, вызываемого стимуляцией пептидергического интернейрона. В то же время увеличение длительности и амплитуды интернейронных ПД наблюдались во время этой процедуры. Стимуляция анального нерва вызывает в нейроне ППа1 постсинаптический ток, состоящий по крайней мере из трех компонентов: двух быстрых с потенциалами реверсии -50 è -70 мВ и долгодлящегося с потенциалом реверсии между -50 è -95 мВ. Сделан вывод, что торможение медленного входящего тока дофамином связано с процессами, происходящими в постсинаптическом нейроне ППа1. Основываясь на гипотезе о том, что берстовая активность нейрона ППа1 является результатом активации пресинаптического неидентифицированного пептидергического интернейрона с постоянной активностью, и на основе представленных данных предполагается, что торможение берстовой активности, вызываемой аппликацией дофамина или же стимуляцией анального нерва, является последствием снижения эффективности синаптической передачи между интернейроном, вызывающим инициацию берстовой активности и собствен-
351
но берстером. Дофамин тормозит синаптическую передачу между интернейроном, инициирующим пейсмекерную активность, и берстовым
нейроном.
Пейсмекер и чувствительность нейронов к веществам в
сверхмалых дозах. Замечательным свойством пейсмекера является его чувствительность к широчайшему спектру воздействий. В эволюционном аспекте эта особенность пейсмекера понятна: впервые появившись у простейших одноклеточных, он был инструментом организации поведения, гарантировавшего выживание. В частности, это свойство пейсмекеров обеспечивает их участие в ответах на биологи- чески активные вещества в сверхмалой концентрации (Бурлакова и др., 2004.).
Ðèñ. 13.15. Изменения пейсмекерной активности под влиянием сверхмалых доз этанола: А — изменение фоновой активности пейсмекера (а) под влиянием этанола в концентрации 10-19М (б); Б — нарушение пластичности нейрона в этанолсодержащей среде: а — развитие привыкания при повторных применениях деполяризационных импульсов силой 0,7 нА, длительностью 3 с в нормальном физрастворе, б — нарушение динамики привыкания через 5 мин. пребывания в физиологическом растворе, содержащем этанол в концентрации 10-15М. Калибровка: 10 мВ, 1 с.
Âэкспериментах мы применяли биологически активные вещества
âнеобычно малых концентрациях, значительно ниже обычно используемых — от 10-15Ì äî 10-28 М (Бурлакова и др., 1987). Результаты показали, что все испытанные вещества: антиоксиданты, нейротранс-
352
миттеры, этанол, ноотропные препараты и др. — в названных концентрациях эффективны и вызывали изменение пейсмекерных ответов на тестовые стимулы (Бурлакова и др., 1986). Следовательно, основной мишенью для действия веществ в столь малых дозах является локус пейсмекерной активности (опыты выполнялись на изолированных нейронах). Некоторые нейроны отвечали включением пейсмекерного механизма или же изменением его состояния (рис. 13.15, А). Это новое состояние клетки проявлялось и в нарушении «обучения» клетки (рис. 13.15, Б). Интересно, что сверхмалые дозы биологически активных веществ вызывают изменения электрической активности и у пара-
меций, следовательно, их поведение в такой среде изменится. Пластичность и пейсмекер. Структурная организация памяти
любого живого существа представлена динамичной организацией и реорганизацией функциональных ансамблей нервных клеток. Нейроны, составляющие системы, имеют индивидуальные свойства, которые характерны только для них. Происхождение индивидуальных свойств нейронов имеет длительную историю, в которой отражаются особенности взаимоотношений данного нейрона с «системой». Динамика этих взаимоотношений определяется свойствами конкретного нейрона и жизненными ситуациями, с которыми сталкивается организм. Реализация имевшегося или приобретение нового опыта выражена активностью нервных клеток: она и есть знание живого существа, высказанное на языке биологических процессов. Если обратиться к современным представлениям о нейронных механизмах памяти, то обнаружим, что локусом пластичности во всех исследованиях является синапс (Martin et al., 2000, Antonov et al., 2001, Соколов и Незлина, 2003). Именно с различными изменениями в области синаптических контактов связывают все наиболее изученные формы обучения: LTP, ассоциативное обучение, привыкание. Присутствие же пейсмекера хотя бы в одном нейроне формирующейся системы позволяет использовать опыт далекого эволюционного прошлого.
В изучении генеза внутриклеточных механизмов памяти весьма полезным и интересным объектом исследования могут быть одноклеточные простейшие. С одной стороны, одна клетка! А с другой, возможности этого существа удивительно многообразны. Поведение одноклеточных сложно, они обучаются, получено неассоциативное и ассоциативное обучение. Поэтому можно предполагать, что пластичность эндогенных осцилляторов одноклеточных является биологической основой обучения.
353
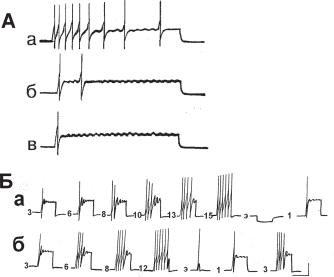
Привыкание. В лаборатории Е. Н. Соколова было показано, что повторные применения внутриклеточных импульсов электрического тока вызывают постепенное изменение реакции нейрона (Пейсмекерный потенциал нейрона, 1975). Явление получило название эндонейрональной пластичности. Предполагалось, что внутриклеточный электрический импульс активизирует пейсмекерный механизм клетки и именно его пластичность проявляется при повторных действиях тока. Но четкие доказательства эндонейронального, а не сетевого происхождения этого феномена можно было получить только на изолированной соме нейрона.
Ðèñ. 13.16. Привыкание и фасилитация пейсмекерных ответов, возникающих при повторном применении внутриклеточных деполяризационных импульсов постоянной силы и длительности. А — привыкание к раздражению силой 0,6 нА, длительностью 5 с, частотой 1 импульс в 30 с. Показаны ответы на 1 (а), 10 (б) и 15 (в) предъявления; Б — фасилитация ответа при повторном действии деполяризационного импульса силой 1,75 нА, 1000 мс, 1 раз в 20 с: а — увеличение ответа и его восстановление экстрастимулом (гиперполяризационным импульсом длительностью 200 мс, силой 0,5 нА; б — фасилитация ответа во второй серии и восстановление ответа деполяризационным импульсом силой 1,75 нА, длительностью 500 мс. Цифры у левого нижнего угла нейрограмм соответствуют номеру стимула. Калибровка: 20 мВ, 1 с.
354
То, что нейроны, входящие в нейронные ансамбли, в зависимости от формы обучения демонстрируют различные формы пластичности, естественно. Эта определяемая обстоятельствами изменчивость их реакций отражает динамику взаимодействия элементов, организующих конкретную нейронную систему. Роль пресинаптического механизма в пластичности нейронных реакций изучалась на самых разных биологических моделях и животных: на срезах мозга, различных препаратах, культуре нервных клеток. Другой реальный пункт пластических изменений нейронных ответов — это сома клетки. Первая задача, для решения которой применили изолированные нейроны, заключалась в изучении изменений, которые получались в ответах нейрона при повторном применении стимула с постоянными физическими параметрами. Результаты таких опытов показали, что, действительно, развиваются явления, которые можно назвать привыканием — постепенно уменьшается исходный ответ, и фасилитацией — постепенно увеличивается исходный ответ (рис. 13.16) (Греченко, Соколов, 1979). Измененную в результате повторного применения стимулов реакцию нейрона можно было восстановить перерывом в стимуляции, причем этот интервал времени, эффективный для восстановления, возрастал по мере выполнения новых серий «обучения». Ключевым параметром для отнесения явления к классу «привыкания» является возможность растормаживания действием экстрараздражителя. В опытах эффективными для восстановления ответов оказались изменение силы, длительности и полярности внутриклеточного тока (рис. 13.16, Б). Необходимо отметить, что не любой экстрастимул оказывался эффективным для полного восстановления ответа, тем не менее всегда удавалось получить хотя бы частичное растормаживание исходной реакции нейрона. Анализируя состав компонентов, участвующих в модификации нейронного ответа, выделяем изменение числа ПД, изменение амплитуды пейсмекерных колебаний и амплитуды ПД (рис. 13.16). Более подробный анализ генеза этих процессов, выполненный при помощи замены ионного состава физиологического раствора, в котором находятся нервные клетки, позволил показать, что, как правило, пластичность достигается участием Са++-зависимых процессов, и лишь иногда — обычно это ПД, меняющие амплитуду, — Na+-зависимых (рис. 13.17). Таким образом, полностью изолированная сома нервной клетки сохраняет пластичность ответов на повторные применения внутриклеточных импульсов тока. Следовательно, процессы, которые обеспечивают пластичность, являются эндогенными по локализации и происхождению.
355
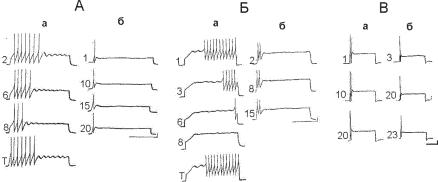
Ðèñ. 13.17. Пейсмекер и кальций. Роль ионов кальция в пейсмекерной активности и привыкании идентифицированного нейрона ЛПа3 к внутриклеточ- ному деполяризационному импульсу силой 2,2 нА, длительностью 5 с: А — нейрон ЛПа3 в состоянии 1 (с актуализацией пейсмекерной активности в нача- ле ответа); Б — тот же самый нейрон в состоянии 2 (с пейсмекерным ответом на поздней фазе действия стимула); В — нейрон в состоянии 3 (с отсутствием пейсмекерной активации). Обозначения: а — электрическая активность нейрона в нормальном физрастворе, б — активность того же самого нейрона в бескальциевом физрастворе. Цифры соответствуют номеру предъявленного стимула. Калибровка: 10 мВ, 1 с.
Ассоциативное обучение. Опыты на изолированных нейронах показали, что исключительное значение при формировании ассоциативных ответов приобретает интервал времени между ассоциируемыми стимулами, в наших опытах — при сочетании микроаппликации нейромедиатора с электрическим внутриклеточным раздражением от 5 мс до 150 мс. При сочетании двух электрических стимулов, наносимых через два независимых внутриклеточных микроэлектрода, локализованных в разных частях сомы, эфективными оказались интервалы от 5 мс до 500 мс (рис. 13.18). Динамика формирования условных ответов на изолированных нейронах, как правило, типична: в каждой следующей серии требуется меньшее количество ассоциированных предъявлений, время сохранения ответа без подкрепления возрастает (Греченко, 1984, 2001). На изолированных нейронах получены клеточные аналоги феномена «напоминания»: после угасания условного ответа однократное предъявление раздражения, похожего на безусловный, приводит к восстановлению условной реакции (ðèñ. 13.18, Á).
356
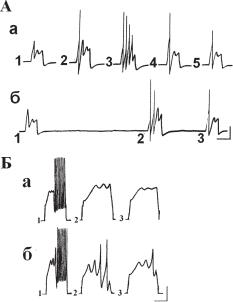
Ðèñ. 13.18. Ассоциативное обучение изолированных нейронов на основе пейсмекерных осцилляций. А — формирование условного ответа при сочетании двух электрических внутриклеточных стимулов на изолированном нейроне Лпа2: а — создание условного ответа при действии условного стимула силой 1,8 нА, длительностью 500 мс (1), подкрепляемого электрическим стимулом силой 2,1 нА, длительностью 500 мс (БС) (2) при сочетании с интервалом между стимулами 50 мс (3). Ответы на изолированные предъявления УС через 1 и 7 мин. (4,5); б — несинхронизированное предъявление УС и БС через 40 мин. после обучения — ответ на УС (1) усиливается (3) после однократного действия БС (2). Калибровка: 10 мВ, 500 мс. Б — ассоциативное обучение неидентифицированного пейсмекерного нейрона: а — результат ассоциативного предъявления УС (1,35 нА, длительность 1000 мс) в сочетании с БС (2,5 нА, длительность 1000 мс) с интервалом между УС и БС 1000 мс (1), после 10 сочетаний (2) и через 5 мин. (3); б — условные ответы после проведения третьей серии сочетаний прежних стимулов. Показано третье сочетание (1) и ответы на изолированные УС через 13 мин. (2) и через 20 мин. (3) после заверешния обучения. Калибровка: 10 мВ, 500 мс.
Обычно результат ассоциативного обучения на клеточном уровне представляют упрощенно: если «условный» стимул вызывает небольшой по амплитуде ответ, то предполагается, что обучение приведет к
357

увеличению его амплитуды, он достигнет порога и будет запускать ПД. Однако такое развитие событий происходит далеко не всегда, невозможно предсказать, как именно нейрон выразит свое участие в процедуре обучения. Возможны различные «превращения» исходных ответов.
Ðèñ. 13.19. Разнообразие способов обучения. А — развитие условного подавления ответа у нейрона Лпа3 при сочетании двух сверхпороговых стимулов. а — ответы нейрона на деполяризационные внутриклеточные раздражения до обучения: 1 — исходный ответ на условный стимул (2,5 нА). 2 — ответ на безусловный стимул (2,6 нА), 3 — ответ на тестовый стимул (2,4 нА), б — ответы после предъявления 8 сочетаний с интервалом между условным и безусловным стимулами 1800 мс, частота предъявлений 1 раз в 45 с. 1 — ответ на первое сочетание, 2 — предъявление восьмой пары стимулов и ответ на тестирующий стимул. Амплитуда ПД полностью не прописана. Калибровка: 20 мВ, 1 с. Б — изменение формы ПД при сочатании сверхпороговых электри- ческих раздражений, предъявленных «cенсорному» нейрону.
Весьма существенна исходная эффективность сочетаемых стимулов, особенно — «условного». В реальной ситуации, когда действует биологически индифферентный стимул, это не означает, что живое существо его не замечает. Оно не знает его смыслого значения, но стимул как раздражитель, поступающий из окружающего мира, им восприниимается. Опыты показали, что результат обучения в таком случае зависит от типа нейрона, от его функции в системе. Если два ассоциированных стимула, достигающих порога генерации ПД, предъявляются гигантскому идентифицированному нейрону ЛПа3, о котором известно, что это командный нейрон, то «условный» ответ заключается
âпостепенном вытормаживании реакции на первый из предъявляемых
âпаре стимулов (условный) (рис. 13.19, А). Тестирующий стимул, предъявляемый через третий внутриклеточный микроэлектрод, показывает, что эта временная депрессия затрагивает только ответ на »условный» cтимул: ни тестирующий, ни «безусловный» никаких изменений не претерпевают (рис. 13.19, А, б). Динамика развития такого ус-
358
ловного подавления ответа типична для процесса ассоциативного обу- чения: в каждой следующей серии скорость развития подавления возрастает, время сохранения условного подавления увеличивается. Особенностью развития условного подавления по сравнению с положительными условными ответами яляется то, что межстимульный интервал может быть необыкновенно большим: в опытах показано, что эффективный интервал между условным и безусловным стимулами от 5 мс до 2000 мс! Учитывая ранее представленные результаты о независимом формировании ответов на нескольких локусах одного и того же нейрона, можно предположить, что такие своеобразные «временные ворота» являются как бы фильтром для подавления всех сверхпороговых активностей, ассоциированных с данным «подкреплением». Положительным же эффектом наделен только подпороговый компонент, подкрепляемый сверхпороговой активностью, поступающей через определенный вход или генерируемой эндогенно, но с задержкой, не превышающей 150 мс. Все остальное — подавляется (Гре- ченко, 2001).
Подавлением ответа на сверхпороговый входной сигнал отвечают крупные идентифицированные нейроны (например, ЛПа3, ППа3), а мелкие клетки левого верхнего квадрата париетального ганглия, которые во многих работах идентифицируются как «сенсорные», в аналогичной ситуации отвечают совершенно иначе. Анализ ответа показал, что по мере нанесения стимулов меняется форма ПД, возникающего на первый в паре стимул («условный»). Изменение ситуации обучения, например, повторные применения стимула, не вызывали такого явления, площадь ПД сокращалась, и все ответы вкладывались один в другой, как матрешки (рис. 13.19, Б)! Увеличение площади ПД, его расширение — специфическая форма выражения обучения, свойственная только ситуации «ассоциативного обучения». В опытах использовалось также предъявление тестирующего стимула, подаваемого через специальный третий внутриклеточный микроэлектрод. Его форма не изме-
нялась на протяжении всего эксперимента (рис. 13.19, Б). Пейсмекер и ионы кальция. Эксперименты показывают, что со-
стояние нейрона и возможность пейсмекерной активности участвовать в его регуляции зависит не только от наличия кальция во внутренней и внеклеточной среде, но и от соотношения его концентраций. В опытах были получены данные, которые показывают, что нейрон в бескальциевой среде фактически не генерирует пейсмекерную активность (рис. 13.17). Тестирование пластичности показало, что такие следовые
359