

15. Потенциал действия сократительныхкардиомиоцитов. Его соотношение с сокращением и фазами возбудимости миокарда.
Клетки миокарда обладают возбудимостью, но им не присущаавтоматия. В период диастолы мембранный потенциал покоя этих клеток стабилен, и его величина выше (80—90 мВ), чем в клетках водителей ритма. Потенциал действия в этих клетках возникает под влиянием возбуждения клеток водителей ритма, которое достигает кардиомиоцитов, вызывая деполяризацию их мембран.
Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной быстрой реполяризации, переходящей в фазу медленной реполяризации (фаза плато), и фазы быстрой конечной реполяризации (рис. 9.8). Фаза быстрой деполяризации создается резким повышением проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Последний, однако, при достижении мембранного потенциала 30—40 мВ инактивируется и в последующем, вплоть до инверсии потенциала (около +30 мВ) и в фазу «плато», ведущее значение имеют кальциевые ионные токи. Деполяризация мембраны вызывает активацию кальциевых каналов, в результате чего возникает дополнительный деполяризирующий входящий кальциевый ток.
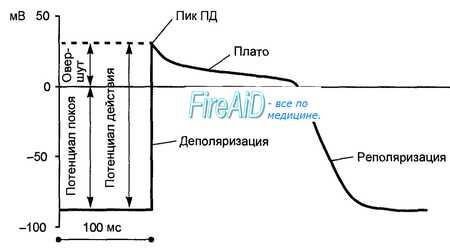
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя. Длительность потенциала действия кардиомиоцитов составляет 300—400 мс, что соответствует длительности сокращения миокарда
16. Электромеханическая связь. Механизм сокращения рабочих кардиомиоцитов. Значение внеклеточногоСа.
Сопряжение возбуждения - сокращения иначе называется электромеханическим сопряжением. Потенциал действия, который запускает процесс, изменение внутриклеточной концентрации Ca2+ [Ca2+]in на его фоне, что запускает механизм сокращения и механограмма, т.е. само сокращение.
Ca2+ существенно важен для сокращения сердца. Удаление Ca2+ из внеклеточной жидкости приводит к уменьшению силы сердечных сокращений и последующей остановке сердца в диастоле. Напротив, увеличение концентрации внеклеточного Ca2+увеличивает силу сердечных сокращений, и очень высокие концентрации Ca2+приводят к остановке сердца в систоле. Свободный внутриклеточный Ca2+ является ионом, отвечающим за сократительную способность миокарда.
|
|
Волна возбуждения в сердечной мышце быстро распространяется вдоль сарколеммы клеток миокарда от клетки к клетке через щелевые контакты. Возбуждение также распространяется внутрь клеток через поперечные трубки, которые инвагинированы в сердечные волокна на полосах Z. Во время плато (фаза 2) потенциала действия повышается проницаемость сарколеммы для Са2+. Са2+ входит в клетку по его электрохимическому градиенту через Са2+-каналы сарколеммы и ее инвагинаций, то есть через мембраны Т-системы.
Считается, что открытие Са2+-каналов происходит в результате фосфорилирования протеинов каналов с помощью циклической аденозинмонофосфат-зависимой протеинкиназы(цАМФ-зависи-мой протеинкиназы). Первоначальным источником внеклеточного Са2+ является интерстициальная жидкость (10-3 М Са2+). Некоторое количество Са2+ может также быть связано с сарколеммой и с гликокаликсом, мукополисахаридом, покрывающим сарколемму. Количество Са2+, который попадает внутрь клетки из внеклеточного пространства, недостаточно для того, чтобы вызвать сокращение миофибрилл. Вошедший внутрь Са2+ («запускающий» Са2+) вызывает высвобождение Са2+ из саркоплазматического ретикулума. [Ca2+]in возрастает от уровня покоя примерно в 10-7 М до уровней от 10-6 до 10-5 во время возбуждения. Затем Са2+ связывается с белком тропонином-С. Кальциево-тропониновый комплекс взаимодействует с тропомиозином, чтобы снять блок с активных участков между актиновыми и миозиновымифиламентами. Это снятие блока позволяет образовываться циклическим поперечным связям между актином и миозином и, следовательно, позволяет миофибриллам сокращаться.
Механизмы, которые повышают [Ca2+]in, увеличивают развиваемую силу сердечных сокращений, а механизмы, которые снижают [Ca2+]in, уменьшают ее.
|
|
В конце систолы приток Са2+ в клетку прекращается и саркоплазматический ретикулум не получает дальнейшей стимуляции для высвобождения Са2+. Фактически саркоплазматический ретикулум начинает активно поглощать Са2+ благодаря кальциевому насосу, который работает за счет энергии аденозинтрифосфата. Работа этого насоса стимулируется фосфоламбаномв результате фосфорилированияфосфоламбанацАМФ-зависимой протеинкиназой. К тому же фосфорилированиетропонина I подавляет связывание Са2+тропонином-С. Этот процесс позволяет тропомиозину снова заблокировать участки взаимодействия актиновых и миозиновых нитей, что приводит к расслаблению (диастоле).
И сокращения, и расслабления сердечной мышцы ускоряются катехоламинами и активацией аденилатциклазы. .. Таким образом, процессы фосфорилирования, вызванные цАМФ-зависимой протеинкиназой, служат для увеличения как скорости сердечных сокращений, так и скорости релаксации.
Митохондрии также поглощают и высвобождают Са2+, но этот процесс протекает слишком медленно.
Са2+, поступающий в клетку для инициирования сокращения, должен быть удален во время диастолы. Это удаление из цитозоляосуществляется прежде всего за счет 3Na+/1Са2+-обменника. Также Са2+ удаляется из клетки электрогенным насосом, который использует АТФ для переноса Са2+ через сарколемму.
Механизм сокращения миокарда существенным образом не отличается то такового в скелетных мышцах. В основе сокращения сердечной мышцы как целостной структуры лежат сокращения сократительного аппарата миокардиальной клетки – миофибрилл. Каждая миофибрилла состоит из параллельно расположенных толстых и тонких нитей особых белков - протофибрилл. На поперечном срезе видно, что каждая толстая протофибрилла окружена 6 тонкимипротофибриллами, а каждая тонкая – 3 толстыми.
Чередование более темных участков, в которых находятся и толстые, и тонкие протофибриллы, с более светлыми участками, в которых находятся только тонкие протофибриллы, создает характерную для сердечной и скелетной мышц поперечную исчерченность. Функциональной единицей сердечной мышцы является саркомер, который представляет собой комплекс одного темного и двух прилегающих к нему светлых дисков.
Толстые протофибриллы состоят из миозиновых нитей. Миозин представляет собой белок, состоящий из двух функциональных частей:
-
хвостовой или фибрильный участок, имеющий вид двух переплетенных ∝-спиралей;
-
глобулярная часть, содержащая две «головки» - G с соединенными с ними легкими цепями L (рисунок.
В состав тонких протофибрилл входит белок актин, который представляет собой двойную спираль (фибриллярный F-актин) образованную глобулами (G-актин) (рисунок 6).
Выступающие «головки» миозина прикрепляются к соседним актиновым нитям, образуя акто-миозиновые мостики. В присутствии ионов Ca2+акто-миозиновый комплекс приобретает способность гидролизовать АТФ с высокой скоростью (рисунок тот же).
Таким образом, в отличие от скелетной мышцы, миокард не способен отвечать тетаническим сокращением на повышение силы раздражителя. Вместе с тем подчинение миокарда закону «все или ничего» не абсолютно. Если раздражать сердечную мышцу импульсами возрастающей частоты, не меняя их силы, то величина сократительного ответа миокарда будет возрастать на каждый последующий стимул. Это явление получило название лестницы. Считается, что механизм возникновения явления лестницы состоит в том, что каждый последующий стимул попадает в фазу повышенной возбудимости мышцы, вызывая тем самым повышенную ответную сократительную реакцию.