

1. Мембранный потенциал, механизмы его формирования. Методы регистрации и параметры. Концентрация ионов калия в клетке почти в 50 раз выше, чем за клеткой, а натрия и хлора больше за клеткой, чем в клетке. Согласно законам диффузии происходит пассивный транспорт ионов по градиенту концентраций: калий стремится выйти из клетки, а натрий- зайти в клетку. Однако мембрана клетки обладает различной проницаемостью для этих ионов. В состоянии покоя мембрана больше проницаема для ионов калия, чем для натрия и анионов. Поэтому в покое преобладает движение ионов калия из клетки над входом в клетку ионов натрия. Калий в клетке находится в связанном состоянии с анионами, которых мембрана не пропускает, и они сосредоточиваются на внутренней поверхности мембраны, обусловливая её отрицательный заряд. А калий, выходя из клетки, сосредоточивается на наружной поверхности мембраны, обеспечивая ей положительный заряд. Таким образом, в состоянии покоя внутренняя поверхность клеточной мембраны заряжена отрицательно, а наружная – положительно. Заряд мембраны клетки, находящейся в состоянии покоя, носит название потенциал покоя или мембранный потенциал клетки. Главная роль в формировании потенциала покоя принадлежит ионам калия. Но если бы потенциал покоя был обусловлен только выходом ионов калия из клетки, то он был бы равен 97,5 мв ( это равновесный потенциал, рассчитанный по формуле Нернста), но такого потенциала не имеет ни одна клетка, что свидетельствует о том, что в состоянии покоя мембрана пропускает в клетку небольшое количество натрия , а также хлора. Следовательно, величина исходного (мембранного) потенциала зависит от того, насколько движение ионов калия из клетки преобладает над входом в клетку ионов натрия.Чем выше соотношение между выходом калия и входом натрия, тем выше мембранный потенциал. Выход же калия зависит от градиента концентраций калия по обе стороны мембраны. И если бы существовал только пассивный транспорт ионов, то он неизбежно привел бы к выравниванию концентраций, а следовательно, и к исчезновению потенциала. Поэтому наряду с пассивным транспортом существует активный транспорт (калий-натриевый насос),осуществляющий движение ионов против градиента концентраций: калия – в клетку, а натрия – из клетки. При этом затрачивается энергия АТФ. Благодаря калий-натриевому насосу восстанавливается градиент концентраций ионов и поддерживается потенциал покоя.Таким образом, прохождение ионов через мембрану, их асимметричное распределение по обе стороны мембраны и связанный с этим электрический потенциал осуществляется посредством 2-х механизмов:1 – свободной диффузией ионов по концентрационному и электрохимическому градиенту; 2 – с помощью натрий- калиевого насоса.У различных клеток мембранный потенциал покоя варьирует от –50 до –90 мВ.Обнаружить МПП можно с помощью второго опыта Гальвани, т.е. зарегистрировать ток покоя между поврежденным и неповрежденным участками мембраны. Для измерения потенциала покоя используют микроэлектродную технику.
2.Потенциал действия, его фазы, механизм формирования. Методы регистрации, параметры. Потенциал действия (ПД) или распространяющееся возбуждение возникает при действии пороговых или сверхпороговых раздражителей, а также в результате суммации подпороговых стимулов, когда достигается критический уровень деполяризации. Возбуждение – это процесс, характеризующийся волнообразными изменениями электропотенциалов на мембране. Выделяют несколько фаз потенциала действия:1)скрытый ( латентный) период или местное возбуждение, когда деполяризация доходит до критического уровня. 2)Деполяризация– дальнейшее снижение исходного потенциала до нуля, а затем перезарядка мембраны ( до + 30 мв.), что регистрируется в виде пика или спайка. 3)Пик ПД ещё называют овершут или инверсия, т.к при этом заряд меняется на противоположный (внутренняя поверхность мембраны заряжается положительно, а наружная – отрицательно). Восходящая часть ПД имеет большую крутизну ( длится от 0,01 до 0,03 сек.), что обусловлено лавинообразным поступлением в клетку ионов натрия. Во время пика блокируются быстрые натриевые каналы и начинается обратный процесс. 4)Реполяризация - восстановление мембранного потенциала вследствие восстановления проницаемости для ионов калия и натрия. Реполяризация или нисходящая часть ПД связана с выходом из клетки ионов калия. Но восстановление исходного потенциала происходит сравнительно медленно, при этом регистрируются следовые потенциалы. 5)Следовая деполяризация или следовой отрицательный потенциал– связан с восстановлением проницаемости для натрия. При этом потенциал близок к критическому уровню. 6)Следовая гиперполяризация или следовой положительный потенциал обусловлен выходом ионов калия из клетки и работой натриевого насоса, осуществляющего активное выведение натрия ,поступившего в клетку в процессе возбуждения.Свойства потенциала действия: 1 – возникает при достижении критического уровня деполяризации; 2- не зависит от силы раздражителя ( если сила подпороговая – ПД не возникает, пороговая или сверхпороговая- ПД возникает не зависимо от силы, подчиняется закону « всё или ничего»); 3 – не способен к суммации; 4 – не обратим; 5 – распространяется без затухания (без декремента). Электроэнцефалография (ЭЭГ) - метод регистрации электрической активности (биопотенциалов) головного мозга.
3.Местный(локальный) ответ, его свойства.Закон «все или ничего». Понятие о деполяризации и гиперполяризации. ЛО возникает при действии раздражителя подпороговой силы (от 0,5 до 0,9 пороговой). Если действует раздражитель ниже 0,5 порогового, не возникает даже местного возбуждения. При местном возбуждении проницаемость мембраны для ионов натрия увеличивается , что приводит к деполяризации мембраны, но она не доходит до того критического уровня, который нужно достичь, чтобы возник потенциал действия. Для большинства тканей он равен ( - 50 мв). Таким образом, критический уровень деполяризации – это уровень деполяризации, при достижении которого начинаетсяПД. Свойства местного возбуждения:1- не подчиняется закону «все или ничего».2- амплитуда ЛО зависит от силы стимула.3- распространяется по мембране затуханием.4- может суммироваться.5- трасформируется в ПД при достижении уровня критической деполяризации. Закон «все или ничего»: При действии подпорогового раздражителя ответной реакции не возникает(«ничего»), при действии порогового или сверхпорогового раздражителя возникает max ответная реакция, величина которой не зависит от силы действующего раздражителя. Т.о. ответ клетки не зависит от силы раздражителя. При нанесении раздражения мембранный потенциал меняется, что связано с изменением проницаемости мембраны. При увеличении проницаемости для ионов натрия ( открытие активационных натриевых каналов) натрий начинает поступать в клетку быстрее и в большем количестве, чем в состоянии покоя, что приводит к снижению исходного потенциала. Это явление ( снижение мембранного потенциала) называется деполяризацией мембраны. Деполяризация характеризует процесс возбуждения. При повышении проницаемости для ионов хлора или калия наблюдается увеличение мембранного потенциала, которое носит название гиперполяризации мембраны; данное явление характерно для торможения.
4. Возбудимость. Порог деполяризации и пороговая сила раздражения как показатели величины возбудимости. Возбудимость – способность тканей в ответ на раздражение давать специфическую ответную реакцию в виде ПД.Порог деполяризации: разность между критическим уровнем деполяризации и величиной исходной поляризации мембраны, зависит от соотношения активированных натриевых и калиевых каналов, порог деполяризации определяет величину пороговой силы раздражения.Возбудимость обратно пропорционально зависит от величины порога деполяризации. Чем выше порог деполяризации тем возбудимость меньше.
5. Фазы возбудимости мембраны нервного и мышечного волокна в процессе возбуждения, рефрактерность. Соотношение с фазами ПД. Фазы возбудимости:1) супернормальная(повышенная) 2)абсолютная рефрактерность – отсутствие возбудимости(раздражитель любой силы не способен вызвать ответной реакции мембраны) 3)относительная рефрактерность(незначительную ответную реакцию может вызвать только сверхпороговый раздражитель) 4)супернормальная(повышенная) 5)субнормальная(пониженная) возбудимость.Соотношение фаз возбудимости с фазами потенциала действия:Во время развития медленной деполяризации возбудимость повышается (мембранный потенциал приближается к критическому уровню деполяризации); во время пика потенциала действия мембрана утрачивает возбудимость, которая постепенно восстанавливается после окончания пика.Период полной невозбудимости получил название «фаза абсолютной рефрактерности». В этот период клетка не отвечает на действии пороговых и сверх пороговых раздражителей.Реполяризация мембраны ведет к реактивации натриевых каналов и снижению калиевой проводимости. Это период так называемой «относительной рефрактерности». В данную фазу возбудимость постепенно нарастает. В дальнейшем фаза относительной рефрактерности сменяется фазой повышенной возбудимости «супернормальности» или экзольтации. В этот момент порог раздражения снижен по сравнению с исходными значениями, поскольку мембранный потенциал ближе к критической величине, чем в состоянии покоя. В таком состоянии клетка может дать ответ на раздражитель, величина которого несколько ниже пороговой.Следовая гиперполяризация, напротив, сопровождается снижением возбудимости (субнормальности).
6. Законы раздражения возбудимых тканей. З-н силы. Понятие порога раздражения. Закон «все или ничего» Раздражение – это процесс воздействия на клетку. Законы раздражения – комплекс законов, которые описывают требования, которым должен удовлетворять раздражитель, чтобы он мог вызвать процесс возбуждения. К ним относятся: полярный закон раздражения, закон силы, времени(длительности действия) и закон крутизны(времени нарастания силы раздражения).Закон силы – чтобы возник ПД,сила стимула должна быть не меньше порогой величины.Существует два проявления этого закона.чем сильнее ( выше пороговой ) раздражитель, тем сильнее ( до определенных пределов) ответная реакция. Другое проявление – закон «всё или ничего»: подпороговый раздражитель не вызывает ответной реакции («ничего»), пороговый и сверхпороговый раздражители дают одинаковую (максимальную) ответную реакцию («всё»). Порог раздражения – минимальная сила, вызывающая ответную реакцию. Закон «все или ничего»: При действии подпорогового раздражителя ответной реакции не возникает(«ничего»), при действии порогового или сверхпорогового раздражителя возникает max ответная реакция, величина которой не зависит от силы действующего раздражителя. Т.о. ответ клетки не зависит от силы раздражителя.
7.Законы раздражения возбудимых тканей. Закон длительности раздражения. Хронаксия. Раздражение – это процесс воздействия на клетку. Законы раздражения – комплекс законов, которые описывают требования, которым должен удовлетворять раздражитель, чтобы он мог вызвать процесс возбуждения. К ним относятся: полярный закон раздражения, закон силы, времени(длительности действия) и закон крутизны(времени нарастания силы раздражения). Закон длительности раздражения(закон времени) – чтобы возник ПД, время действия стимула должно быть не меньше пороговой величины. Хронаксия(в схеме зависимости между силой и временем действия раздражителя) – min длительность раздражающего импульса при его силе, равной удвоенной реобазе, необходимая для возникновения ПД.
8. Законы раздражения возбудимых тканей. Закон крутизны изменения раздражителя. Аккомодация Раздражение – это процесс воздействия на клетку. Законы раздражения – комплекс законов, которые описывают требования, которым должен удовлетворять раздражитель, чтобы он мог вызвать процесс возбуждения. К ним относятся: полярный закон раздражения, закон силы, времени(длительности действия) и закон крутизны(времени нарастания силы раздражения). Закон крутизны – чтобы возник ПД, крутизна стимула должна быть не меньше пороговой величины. Аккомодация – приспособление ткани к длительно действующему и медленно нарастающему по силе раздражителю.Закон аккомодации – отражает зависимость ответной реакции от скорости нарастания силы раздражителя до определенной (пороговой) величины. При воздействии медленно нарастающих по силе раздражителей увеличивается порог возбудимости, что обусловлено повышением критического уровня деполяризации и инактивацией( закрытием ) быстрых натриевых каналов. При медленном нарастании силы раздражителя до пороговой величины натриевые каналы закрываются раньше, чем потенциал достигнет критического уровня. В данном случае развивается стойкая деполяризация мембраны. Стойкая деполяризация сопровождается низкой возбудимостью, так как в этом случае не достигается критический уровень деполяризации, а следовательно, не возникает возбуждения.
9. Полярный закон раздражения. Полярный закон – при внеклеточном приложении прямоугольного импульса постоянного тока возбуждение возникает при замыкании цепи под катодом, а при размыкании цепи – под анодом. 1)При замыкании цепи электроды приобретают заряды. При этом к катоду, имеющему «-» заряд, притягиваются «+» ионы как с поверхности мембраны, так и изнутри. В результате заряд мембране деполяризуется и при достаточной силе импульса раздражения развивается ПД.В это время к аноду, имеющему «+» заряд, притягиваются «-» ионы, что приводит к развитию гиперполяризации мембраны. 2)При размыкании цепи электроды теряют свои заряды и ионы, которые находились в поле их притяжения, возвращаются к мембране. Поэтому под катодом развивается гиперполяризация, а под анодом – деполяризация, а при достаточной силе импульса –ПД.
10. Изменения возбудимости ткани под влиянием постоянного тока (физиологическийэлектротон). Катодическая депрессия. Физиологическийэлектротон – пассивное изменение возбудимости мембраны под катодом и анодом при воздействии на нее постоянного тока подпороговой силы. При этом под катодом развиваетсякатэлектротон, проявляющийся увеличением возбудимости, а под анодом – анэлектротон, хар-ся снижением возбудимости. Катодическая депрессия по Вериго: 1- если постоянный ток действует на мембрану длительное время, то повышенная возбудимость под катодом изменяется на снижение возбудимости.2-в основе этого явления лежит явление аккомодации ткани, т.к. постоянной ток можно представить как ток с бесконечно малой крутизной нарастания.
11.Физиологические свойства скелетных мышц, их иннервация. Изотонический и изометрический режимы сокращения. Свойства: возбудимость,проводимость,сократимость + эластичность(физическое св-во). Иннервация осуществляется соматической НС. К каждому мышечному волокну подходит аксоннаятерминаль и образует с ним нервно-мышесный синапс. Адекватный раздражитель – только нервный импульс. Во время сокращении мышцы может меняться её длина, а напряжение оставаться на постоянном уровне, такой режим сокращения называется изотоническим. Другой режим сокращения –изометрический, когда при нарастании напряжения длина мышцы не изменяется. Чаще всего наблюдается ауксотонический режим сокращения, характеризующийся изменением длины и напряжения мышцы.
12.Одиночное мышечное сокращение, его фазы, продолжительность. Нейромоторная единица. Понятие о быстрых и медленных мышцах. Одиночное сокращение- возникает при действии одиночного стимула, приложенного к двигательному нерву – непрямое раздражение,или непосредственно к мышце – прямое раздражение. Сократительные свойства обусловлены структурными элементами мышц - миофибриллами, которые в свою очередь представлены сократительными белками: актином (тонкие нити) и миозином (толстые нити). В основе сокращения лежит процесс скольжения нитей актина вдоль миозина (образование акто-миозинового комплекса), в результате чего мышца укорачивается. Чтобы запустить процесс сокращения, нужен сигнал о сокращении, который в естественных условиях поступает от мотонейронов спинного мозга, который вместе с эфферентными нервными волокнами и иннервируемые им мышечными волокнами составляют двигательную или нейромоторную единицу. Следовательно, сокращению мышцы предшествует её возбуждение. 3 механические фазы (периоды) : 1- латентный ( от начала действия раздражителя до начала сокращения), длится 0,01 сек.; 2- укорочение (0,04 сек.); 3 – расслабление (0,05сек.). Каждой механической фазе соответствуют определенные электрические явления на мембране мышечного волокна: латентный период совпадает с возбуждением мембраны (деполяризация и пик потенциала действия); в период укорочения мышцы мембрана реполяризуется , во время максимального укорочения регистрируется следовой отрицательный потенциал (следовая деполяризация), а в период расслабления на мембране восстанавливается исходный (мембранный) потенциал. Все фазы мышечного сокращения нуждаются в энергии АТФ. При недостатке АТФ (действие ядов, утомление и т.д.) возможно развитие контрактуры мышц – состояние длительного сокращения без расслабления (например, «трупное окоченение» после смерти). В процессе возбуждения и сокращения мышцы меняется её возбудимость. В латентный период сокращения мышца абсолютно рефрактерна (это связано с пиком ПД), в период укорочения – возбудимость восстанавливается (относительная рефрактерность), достигая максимума в момент максимального укорочения (связано со следовой деполяризацией мембраны), во время расслабления возбудимость нормальная (исходная), как в состоянии покоя. Классификация мышечных волокон: 1-медленные фазические волокна окислительного типа. Отличаются большим содержанием миоглобина и митохондрий. Имеют красный цвет, медленно утомляются. Входят в состав мышц, поддерживающих позу.2-быстрые фазические волокна окислительного типа. Содержат много митохондрий. Способны синтезировать АТ путем окислительного фосфорилирования. Выполняют быстрые сокращения. Утомляются медленно.3-быстрые фазические с гликолитическим типом окисления. Содержат мало митохондрий, АТФ образуется за счет гликолиза. Миоглобина нет(белый цвет). Быстро сокращаются и быстро утомляются.4-тонические волокна. Двигательный аксон образует множество синапсов. Медленно сокращаются и медленно расслабляются. Входят в состав наружных мышц глаза.
13.Суммация одиночных мышечных сокращений. Формирование гладкого и зубчатого тетануса. Оптиум и пессиум частоты раздражения. Скелетная мышца может реагировать на дополнительный сигнал о сокращении, когда ещё не пройден весь цикл мышечного сокращения., то есть она способна к суммированному (тетаническому) сокращению. В зависимости от частоты поступающих сигналов ( а именно от того, в какую фазу одиночного мышечного сокращения застаёт мышцу новый сигнал о сокращении) различают зубчатый и гладкий тетанус. Если очередной сигнал застаёт мышцу в период расслабления – возникает зубчатый тетанус, в период укорочения – гладкий тетанус. В естественных условиях режим сокращения одной нейро-моторной единицы – зубчатый тетанус, но сокращение целой мышцы – слитное (т.е. гладкий тетанус), так как наблюдается асинхронность разрядов мотонейронов. Суммарная амплитуда сокращения зависит от состояния возбудимости мышцы в момент, когда приходит очередной сигнал о сокращении. Оптимум соответствует частоте импульсов, совпадающих с периодом её повышенной возбудимости (экзальтации). Дальнейшее увеличение частоты импульсов застаёт мышцу в состоянии относительной и абсолютной рефрактерности, что приводит к пессимуму.
14. Структура нервно-мышечного синапса. Медиатор. Механизм передачи возбуждения. Синапс – это специальная структура для передачи импульсов от клетки к клетке. Каждый синапс имеет пресинаптическую и постсинаптическую мембраны и синаптическую щель между ними. Пресинаптическая мембрана покрывает окончание аксона мотонейрона. Через нее в синаптическую щель поступает медиатор, который затем взаимодействует с рецепторами постсинаптической мембраны. Медиатор – вещество, выделяющееся из пресинаптического окончания и изменяющее мембранный потенциал постсинаптической мембраны, увеличивая ее проницаемость для опреденных ионов, что приводит к де или гиперполяризации последней. Механизм передачи возбуждения в синапсе состоит из 3-х этапов:1 – ПД, возникший в пресинаптической мембране, повышает ее проницаемость для ионов кальция, который входит в нервное окончание и освобождает медиатор ацетилхолин из пузырьков.2 – Ацетилхолин диффундирует в синаптическую щель, избыток медиатора разрушает фермент ацетилхолинэстераза.3 – Ацетилхолин взаимодействует с холинорецепторами постсинаптической мембраны, в результате чего открываются натриевые каналы для входа натрия внутрь клетки – развивается потенциал действия, который распространяется вдоль мышечного волокна, вызывая его сокращение. Нервно-мышечный синапс имеет следующие свойства: 1.одностороннее проведение возбуждения, 2.синаптическую задержку (на выход медиатора и его диффузию), 3.низкую лабильность (до 100 имп/сек), 4.быструю утомляемость, 5.высокую чувствительность к химическим веществам, лекарственным препаратам, токсинам (что может ускорять или тормозить передачу импульсов от нерва к мышце).
15.Внутриклеточный механизм сокращения и расслабления мышечного волокна. Сопряжение возбуждения мембраны и сокращения(электромеханическая связь) Механизм сокращения объясняется теорией скольжения: тонкие актиновые нити скользят вдоль толстых миозиновых за счет образования мостиков, в результате уменьшается размер саркомера и длина мышечного волокна. 1. Источником энергии для мышечного сокращения являются молекулы АТФ.2. Гидролиз АТФ катализируется при мышечном сокращении миозином, обладающим ферментативной активностью.3. Пусковым механизмом мышечного сокращения является повышение концентрации ионов кальция в саркоплазме миоцитов, вызываемое нервным двигательным импульсом.4. Во время мышечного сокращения между тонкими и толстыми нитями миофибрилл возникают поперечные мостики или спайки.5. Во время мышечного сокращения происходит скольжение тонких нитей вдоль толстых, что приводит к укорочению миофибрилл и всего мышечного волокна в целом. Электромеханическое сопряжение:1-кальций соединяется с тропонином на актиновых волокнах.2-в результате тропомиозин освобождает активные поверхности актина.3-к этим поверхностям «приклеиваются» головки миозиновых мостиков и образуется акто-миозиновый комплекс-в результате происходит расщепление молекулы АТФ и выделение энергии. Особенности механизма скольжения актиновых и миозиновых волокон:1-кальций в цитоплазме связывается с кальмодулином и активирует протеинкиназу.2-протеинкиназа фосфорилирует головку миозина.3-образуются мостики между миозином и актином.4-уменьшение концентрации кальция в миоплазме вызывает дефосфорилирование головки миозина-мостики распадаются.5-мышца расслабляется Механизм расслабления мышцы:1-при определенной концентрации кальция в цитоплазме включается кальевый насос в СПР.2-кальциевый насос с затратой энергии АТФ «закачивает» кальций обратно в СПР.3-мостики расщепляется,т.к. активные поверхности актина вновь блокируются тропомиозином
16.Сила и работа мышц. Закон средних нагрузок. Эргография Сила мышцы определяется максимальным грузом, который мышца может поднять. Мышцы способны совершать работу. Работа мышц определяется произведением величины поднятого груза на высоту подъема. Максимальная работа производится при средних величинах нагрузок. Лабильность мышцы равна 200–300 Гц. Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках. Работа мышц измеряется произведением поднятого груза на величину укорочения мышцы. Между грузом, который поднимает мышца, и выполняемой ею работой существует следующая закономерность. Внешняя работа мышцы равна нулю, если мышца сокращается без нагрузки. По мере увеличения груза работа сначала увеличивается, а затем постепенно падает. Наибольшую работу мышца совершает при некоторых средних нагрузках. Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов. Эргография:1-метод регистрации работы, выполненной мышцами.2-на кривой (эргограмме) видны признаки утомления: снижение амплитуды сокращения, удлинение фазы расслабления, уменьшение частоты сокращения.
17.Утомление,факторы, влияющие на скорость его развития. Природа и локализация утомления в нервно-мышечном препарате и целом организме. Активный отдых по Сеченеву. Механизмы утомления мышцы можно подразделить на 2 категории: 1.местные причины, связанные в основном с двумя факторами: а) накоплением продуктов обмена и ионов калия в межклеточном веществе, что угнетает способность мембраны генерировать потенциал действия; б) истощением энергетических запасов , связанных со снижением ресинтеза АТФ в результате уменьшения запасов гликогена, креатинфосфата , недостатка кислорода и т.д. 2.центральные механизмы, связанные с утомлением нервных центров, регулирующих сокращение тех или иных мышц. Утомление нервных центров наступает в результате утомления синапсов. В целостном организме быстрее наступает утомление нервных центров, чем самой мышцы. Утомление центрального происхождения легче снять, чем утомление , связанное с местными факторами. Для этого достаточно переключить деятельность на другую группу мышц ( или на другой вид работы), а следовательно, на другие нервные центры. Другими словами, надо создать новый доминантный очаг возбуждения, который затормозит раннее работающие центры и создаст условия для более быстрого их восстановления. На основе этих механизмов И.М.Сеченов выдвинул идею активного отдыха. Утомление в нервно-мышечном препарате:1-процесс временного снижения работоспособности мышцы.2-возникает в связи с накоплением недоокисленных продуктов и уменьшении pH, уменьшением энергетических запасов(АТФ) в мышечном волокне или уменьшением медиатора в нервно-мышечном синапсе.3-в нервно-мышечном препарате утомление раньше развивается в синапсе!Утомление целого организма развивается в начале в нервных центрах –это защитный механизм. При этом в мышцах остается «аварийный» запас энергии. Доказательство- опыты Сеченова с активным отдыхом. Он доказал, что наиболее быстрое восстановление работоспособности руки после утомительной работы наступает не при полном покое обеих рук, а при работе другой, не работавшей руки.
18.Структурная и функциональная характеристика гладких мышц. Более примитивной и малодифференцированной является гладкомышечная ткань, выстилающая стенки сосудов, полых внутренних органов. Несмотря на клеточное строение, её можно отнести к функциональному синцитию: контакты между клетками очень плотные («нексусы») с низким сопротивлением, что способствует беспрепятственному распространению возбуждения с одной группы клеток на другую. Поэтому для гладких мышц характерны медленные длительные тонические сокращения, что необходимо для поддержания тонуса сосудов, органов желудочно-кишечного тракта и др. Кроме того, гладкие мышцы обладают пластичностью –способностью сохранять приданную растяжением длину без изменения напряжения, что имеет важное значение для полых органов. В отличие от скелетных мышц, сокращение которых инициируются сигналами из ЦНС, гладкие мышцы способны к автоматии, т.е. к самовозбуждению, что обусловлено отсутствием стабильного потенциала покоя и наличием медленной деполяризации мембраны, достигающей критического уровня самопроизвольно. Возникнув в одной группе клеток, возбуждение распространяется через «нексусы» (выполняющих роль электрических синапсов) на другие участки, поддерживая тем самым тоническое сокращение (медленное сокращение и расслабление при малых затратах энергии). В формировании потенциала действия гладких мышц принимают участие не быстрые натриевые каналы ( для гладких мышц не характерен «пик» потенциала действия), а медленные натрий-кальциевые каналы, приводяшие к малой крутизне ПД. Кальций, поступающий в мышечное волокно при возбуждении, играет ведущую роль в осуществлении связи между возбуждением и сокращением (электромеханическом сопряжении). Помимо способности к автоматии, которой обладает определенная группа миоцитов – «пейсмеккеров», гладкие мышцы способны возбуждаться при растяжении ( в этом заключается миогенный механизм поддержания тонуса гладких мышц) и под действием медиаторов вегетативной нервной системы.
19.Проведение возбуждения по нервному волокну. Распространение возбуждения по безмиелиновым и миелиновым нервным волокнам. Классификация нервных волокон Нервные волокна — аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских клеток в периферических нервах. Нервные волокна подразделяют на 2 типа — безмиелиновые и миелиновые. Основная функция нервных волокон — проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна и существенно зависит от диаметра нервных волокон.Скорость проведения в миелиновых волокнах выше, чем в безмиелиновых. Безмиелиновые нервные волокна. В покое мембрана аксона (осевого цилиндра) поляризована — положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется, и наружная поверхность мембраны приобретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны. Такое проведение известно как электротоническое, а проведение ПД — своего рода «эстафета», в которой каждый участок мембраны является сначала раздражаемым, а затем раздражающим.Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5–2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения ПД. Миелиновое нервное волокно состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания собственной плазматической мембраны. Миелин прерывается через регулярные промежутки (от 0,2 до 2 мм) концентрической щелью шириной около 1 мкм, это узлы, или перехваты Ранвье. Таким образом, межузловые сегменты аксона, расположенные между соседними перехватами Ранвье, содержат миелин — электрический изолятор, не позволяющий проходить через него локальным токам, поэтому ПД возникают только в перехватах Ранвье. Другими словами, ПД перемещается вдоль нервного волокна скачками, от одного перехвата Ранвье к другому перехвату (скачкообразное проведение).В физиологических условиях ПД движутся в одном направлении от места раздражения. ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения.
20.Рефлекс как основной и универсальный акт нервной деятельности, структура соматического и вегетативного рефлекса .Понятие о рецептивном поле рефлекса. Обратная афферентация. Рефлекс – это закономерная, приспособительная реакция организма в ответ на изменившиеся условия внешней или внутренней среды с участием ЦНС. Рефлекс начинается с рецептивного поля. У каждого рефлекса свое рецептивное поле и свой нервный центр. Морфологической основой рефлекса является рефлекторная дуга – комплекс специфически организованных нервных элементов, взаимодействие которых необходимо для осуществления рефлекторного акта. Любая рефлекторная дуга включает З функциональных отдела или звена: а) афферентное ( чувствительное ) звено, обеспечивающее доставку информации в мозг; 2- центральное звено, где происходит обработка ( анализ и синтез) полученной информации и формирование программы ответной реакции ; 3 – эфферентное (двигательное) звено, благодаря которому сигналы из ЦНС поступают к исполнительным органам, обеспечивая реализацию ответной реакции и получение определенного результата. По современным представлениям рефлекторная дуга включает обратную связь, несущую информацию о полученном результате от исполнительных органов в ЦНС, что позволяет вовремя корректировать ответные реакции. Таким образом рефлекторная дуга превращается в рефлекторное кольцо. Афферентное звено рефлекса начинается с рецептора-специализированной структуры, воспринимающей определенный вид воздействия внешней или внутренней среды. Рецептором может быть окончание дендрита 1-го чувствительного нейрона, тело которого находится в чувствительном спинномозговом узле, либо специальная клетка ( например, фоторецепторы сетчатки глаза , волосковые клетки кортиева органа и др.). Рецепторы могут располагаться на поверхности тела (экстерорецепторы), во внутренней среде (интерорецепторы) или в органах движения ( проприорецепторы). В зависимости от природы раздражителя, действующего на соответствующий рецептор, различают механо-, хемо-, термо- и другие рецепторы. Совокупность рецепторов, раздражение которых вызывает соответствующий рефлекс, называется рецептивное поле или рефлексогенная зона рефлекса. Каждый безусловный рефлекс можно вызвать с определенного рецептивного поля. Например, рецептивным полем слюноотделительного рефлекса является ротовая полость, кашлевого рефлекса- воздухоносные пути и т.д. Под действием раздражителя в рецепторе возникает рецепторный потенциал, имеющий свойства местного возбуждения. При достижении критического уровня он становится потенциалом действия. Чаще всего потенциал действия возникает в теле чувствительного нейрона ( в области аксонного холмика) и по его аксону поступает в центральное звено, где через вставочный нейрон ( а может и без него) переключается на эфферентное звено рефлекса, включающее эфферентный нейрон ( двигательный или вегетативный), эфферентные нервные волокна и исполнительный орган (эффектор). Таким образом, можно проследить следующую последовательность проведения возбуждения в структурах рефлекторной дуги: рецептор-дендрит чувствительного нейрона- тело чувствительного нейрона- аксон чувствительного нейрона- вставочный нейрон- эфферентный нейрон- эфферентное волокно – эффектор. Время от начала раздражения до ответной реакции эффектора называется временем рефлекса.Оно определяется временем проведения возбуждения в афферентном, центральном и эфферентном звене рефлекса и зависит от типа нервных волокон, образующих афферентные и эфферентные пути, от количества переключений в центральном звене, а также от силы раздражителя и от возбудимости нервных элементов рефлекторной дуги.. Главным фактором, влияющим на время рефлекса, является количество синапсов в рефлекторной дуге. Чем сложнее рефлекс и чем больше синапсов, участвующих в проведении возбуждения, тем длительнее время рефлекса. Это связано с задержкой возбуждения в синапсах.
21.Строение и функциональные свойства синапсов ЦНС.Передача возбуждения в них. Возбуждение с нервного волокна на нервную, мышечную и железистую клетку передается посредством специального структурного образования — синапса.Синапс — структура, обеспечивающая проведение сигнала от одной клетки к другой. Строение синапса. Синапсы состоят из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 1).
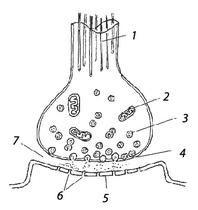
Рис. 1. Строение синапса: 1 — микротрубочки; 2 — митохондрии; 3 — синаптические пузырьки с медиатором; 4 — пресинаптическая мембрана; 5 — постсинаптическая мембрана; 6 — рецепторы; 7 -синаптическая щель
Пресинаптическая мембрана покрывает расширенное нервное окончание, которое представляет собой нейросекреторный аппарат. В пресинаптической части находятся пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в гранулах (пузырьках).
Постсинаптическая мембрана - утолщенная часть мембраны клетки, с которой контактирует пресинаптическая мембрана. Она имеет ионные каналы и способна к генерации потенциала действия. Кроме того, на ней расположены специальные белковые структуры — рецепторы, воспринимающие действие медиаторов.
Синаптическая щель представляет собой пространство между пресинаптической и постсинаптической мембранами, заполненное жидкостью, близкой по составу к плазме крови.
Функциональная классификация синапсов:
1) возбуждающие синапсы;
2) тормозящие синапсы.
3. По механизмам передачи возбуждения в синапсах:
1) химические;
2) электрические.
Особенность химических синапсов заключается в том, что передача возбуждения осуществляется при помощи особой группы химических веществ – медиаторов.
Различают несколько видов химических синапсов:
1) холинэргические. В них происходит передача возбуждения при помощи ацетилхолина;
2) адренэргические. В них происходит передача возбуждения при помощи трех катехоламинов;
3) дофаминэргические. В них происходит передача возбуждения при помощи дофамина;
4) гистаминэргические. В них происходит передача возбуждения при помощи гистамина;
5) ГАМКэргические. В них происходит передача возбуждения при помощи гаммааминомасляной кислоты, т. е. развивается процесс торможения.
Особенность электрических синапсов заключается в том, что передача возбуждения осуществляется при помощи электрического тока. Таких синапсов в организме обнаружено мало.Синапсы имеют ряд физиологических свойств:
1) клапанное свойство синапсов, т. е. способность передавать возбуждение только в одном направлении с пресинаптической мембраны на постсинаптическую;
2) свойство синаптической задержки, связанное с тем, что скорость передачи возбуждения снижается;
3) свойство потенциации (каждый последующий импульс будет проводиться с меньшей постсинаптической задержкой). Это связано с тем, что на пресинаптической и постсинаптической мембране остается медиатор от проведения предыдущего импульса;
4) низкая лабильность синапса (100–150 имульсов в секунду).Передача возбуждения в синапсе.

В рецепторных участках постсинаптической мембраны медиатор взаимодействует с белково-липидными комплексами в результате чего происходит повышение ее проницаемости для ионов Na+, К+, CI-. В этом процессе большая роль принадлежит ферментам (аденилатциклаза). Это приводит к деполяризации постсинаптичской мембраны и возникает возбуждающий постсинаптический потенциал (ВПСП). Когда он достигает критического уровня образуется потенциал действия (ацетилхолин). Медиатор в тормозных синапсах увеличивает проницаемость постсинаптической мембраны только для ионов К+ и CI-. В этом случае происходит гиперполяризация постсинаптической мембраны и генерируется тормозной постсинаптический потенциал (ТПСП).
22.Медиаторы в цнс.Их классификация
В зависимости от химической природы их можно разделить на 4 группы:

По действию медиаторы можно разделить на ионотропных и метаболотропни. Ионотропных медиаторы после взаимодействия с циторецепторамы постсинаптической мембраны изменяют проницаемость ионных каналов. Метаболотропни медиаторы постсинаптическую действие проявляют путем активации специфических ферментов мембраны. Вследствие этого в мембране или (чаще) в цитоплазме клетки активируются так называемые вторичные посредники (вторичные мессенджеры), которые в свою очередь запускают каскады внутриклеточных процессов, тем самым влияя на функции клеток.
По функциональным свойствам медиаторы ЦНС делятся на возбуждающие, тормозные и модулирующие. Возбуждающими медиаторами могут быть различные вещества, которые вызывают деполяризацию постсинаптической мембраны. Важнейшее значение имеют производные глутаминовой кислоты (глутамата), субстанция Р. Некоторые центральные нейроны имеют холинорецепторы, т.е. содержат на постсинаптической мембране рецепторы, которые реагируют с холинового соединениями, например, ацетилхолин в клетках Реншоу .. возбуждающими медиаторами могут быть также моноамины (норадреналин, дофамин , серотонин). € основания считать, что тип медиатора, который образуется в синапсе, обусловлен не только свойствами окончания, но и общим направлением биохимических процессов во всем нейроне.
Природа тормозного медиатора до конца не установлена. Полагают, что в синапсах различных нервных структур эту функцию могут выполнять аминокислоты - глицин и ГАМК.
23.Передача возбуждении в синапсах ЦНС.Возбуждающие синапсы,ВПСП,его ионные механизмы Потенциал действия, пришедший по пресинаптическому волокну к синапсу, вызывает деполяризацию мембраны, которая включает кальциевый насос, и ионы кальция поступают в синапс. Попадая в цитоплазму синаптического окончания, кальций связывается с белками оболочки синаптических везикул (пузырьков, в которых хранятся медиаторы), что приводит к выделению медиаторов в синаптическую щель, которая отделяет мембрану одного нейрона от мембраны другого. Так возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в импульс химический, т. е. каждое возбуждение нейрона сопровождается выбросом в окончании его аксона порции биологически активного вещества – медиатора. Далее молекулы медиатора связываются со специальными белковыми молекулами, которые находятся на мембране другого нейрона. Эти молекулы называются рецепторами. Рецепторы устроены уникально и связывают только один тип молекул (которые подходят как «ключ к замку»). Рецепторы – белковые структуры, которые являются интегральными белками плазматической мембраны. Они синтезируются в рибосомах эндоплазматического ретикулюма клетки, затем встраиваются в мембраны. Функциональная активность синапса зависит от количества рецепторов, а также от их сродства к медиатору (лиганду). Искусственно созданные лиганды, обладающие высоким специфическим (т. е. действующим только на определенный тип либо подтип рецептора) сродством к рецепторам, способны вызывать такой же функциональный ответ клетки, как и нативные медиаторы. Рецептор состоит из двух частей. Одну можно назвать «узнающим центром», другую – «ионным каналом». Если молекулы медиатора заняли определенные места (узнающий центр) на молекуле рецептора, то ионный канал открывается и ионы начинают входить в клетку (ионы натрия) или выходить (ионы калия) из клетки. Другими словами, через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране. Этот потенциал получил название постсинаптического потенциала. В зависимости от характера открытых ионных каналов возникает возбудительный (открываются каналы для ионов натрия и калия) постсинаптсинаптический потенциала (ВПСП) или тормозной (открываются каналы для ионов хлора) постсинаптический потенциал (ТПСП). На мембране одного нейрона могут одновременно находиться два вида синапсов: тормозные и возбудительные. Все определяется устройством ионного канала мембраны. Мембрана возбудительных синапсов пропускает как ионы натрия, так и ионы калия. В этом случае мембрана нейрона деполяризуется. Мембрана тормозных синапсов пропускает только ионы хлора и гиперполяризуется. Очевидно, что если нейрон заторможен, потенциал мембраны увеличивается (гиперполяризация). Таким образом, нейрон благодаря воздействию через соответствующие синапсы может возбудиться или прекратить возбуждение, затормозиться. Все эти события происходят на соме и многочисленных отростках дендрита нейрона (на последних находится до нескольких тысяч тормозных и возбудительных синапсов).
После каждого проведения импульса медиаторы разрушаются специфическими ферментами либо происходит их обратный захват в пресинаптическое окончание. В синапсах существует широко распространенное в физиологии явление обратной связи: медиатор из синаптической щели может взаимодействовать с рецепторами на пресинаптической мембране (ауторецепторами), что приводит к прекращению выброса медиатора. Активность синапса может модулироваться действием модулирующих нейромедиаторов, рецепторы которых располагаются на аксоне или даже дендритах и теле синапса. Если через синапс проходит много импульсов, то рецепторы могут уменьшить свою чувствительность к нейромедиатору (это свойство рецепторов обеспечивает процессы адаптации на нейрональном уровне). Важно помнить, что между химической природой синапса и знаком его синаптического действия (возбуждающий или тормозной) нет однозначной зависимости: один и тот же медиатор может оказывать как тормозное, так и возбуждающее действие. Знак синаптического действия определяется свойствами постсинаптической мембраны, т. е. составом рецепторов, которые могут разным образом реагировать с медиатором и контролировать проводимость разных ионных каналов.