

6.6.1. Первичная структура рнк
Под первичной структурой РНК понимают последовательность рибонуклеотидов, связанных 3–5-фосфодиэфирными связями (похожа на ДНК).
6.6.2. Вторичная структура рнк
Макромолекулы большинства природных РНК построены из одной полирибонуклеотидной цепи. Основной элемент их вторичной структуры – сравнительно короткие двойные спирали, образованные комплементарными участками одной и той же цепи и перемежающиеся ее однотяжевыми сегментами. Полирибонуклеотидные цепи в таких двуспиральных структурах антипараллельны, а сами двойные спирали, находящиеся в А-форме, не идеальны: в них имеются дефекты в виде неспаренных остатков или не вписывающихся в двойную спираль однотяжевых петель (шпильки).
Стабильность двутяжевых районов поддерживается комплементарными (А–У и Г–Ц) и межплоскостными (стэкинг) взаимодействиями оснований. В однотяжевых участках наблюдаются сильные стэкинг-взаимодействия оснований, вследствие чего они стремятся принять конформацию однотяжевой спирали.
6.6.3. Третичная структура рнк
В физиологических условиях однотяжевые РНК характеризуются компактной и упорядоченной третичной структурой, которая возникает за счет взаимодействия шпилькообразных элементов их вторичной структуры. Транспортные РНК – единственные представители природных полирибонуклеотидов, которые удалось закристаллизовать и изучить методом рентгеноструктурного анализа. Поэтому представления о принципах организации структуры РНК практически полностью основаны на данных о пространственной структуре тРНК.
Рассмотрим эти принципы.
1. Элементы вторичной структуры РНК располагаются друг относительно друга так, чтобы обеспечить максимальный стэкинг оснований в макромолекуле в целом.
2. Контакты между отдельными элементами вторичной структуры осуществляются за счет нескольких типов так называемых «третичных» внутримолекулярных взаимодействий:
а) за счет образования дополнительных, зачастую неуотсон-криковских (например, Г–У) пар оснований между нуклеотидными остатками удаленных друг от друга (в первичной и вторичной структурах) однотяжевых участков и триплетов оснований между нуклеотидными остатками однотяжевых и двутяжевых элементов:
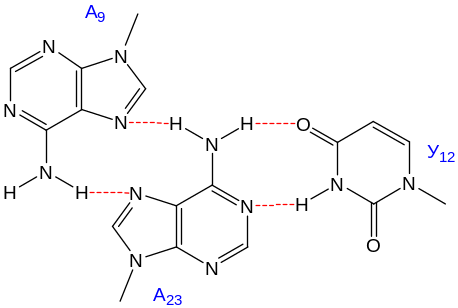
(в тРНКфен дрожжей; уотсон-криковская пара – А23–У12);
б) за счет дополнительных («третичных») стэкинг-взаимодействий после интеркаляции (от англ. intercalation – внедрение) оснований одного участка между двумя соседними основаниями другого однотяжевого участка;
в) за счет образования дополнительных водородных связей между 2-ОН-группами остатков рибозы и основаниями, а также другими группами сахарофосфатного остова.
3. Третичная структура РНК стабилизирована ионами двухвалентных металлов, которые связываются не только с фосфатными группами, но и с основаниями.
6.6.4. Классы рнк
По составу, структуре и функциям различают транспортные (тРНК), рибосомальные (рРНК), информационные (иРНК, или РНК-посредники, мессенджер-РНК, от англ. messenger – курьер, посыльный, мРНК, матричные РНК), ядерные (яРНК, или гетерогенные ядерные РНК – гяРНК).
Транспортные РНК
Основной функцией этих РНК является перенос аминокислот к местам синтеза белка – к рибосомам.
Каждая индивидуальная тРНК способная переносить единственную аминокислоту. Конкретные тРНК называют по имени этой аминокислоты. Например, аланиновая тРНК, или тРНКала. Если одна и та же аминокислота переносится несколькими тРНК, то последние называют изоакцепторными и нумеруют (например, тРНКвал1, тРНКвал2).
По сравнению с другими классами РНК, тРНК богаты минорными нуклеотидными остатками и содержат нуклеозиды и нуклеотиды своеобразного строения:
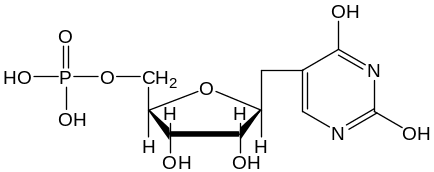 Псевдоуридиловая
кислота ()
Псевдоуридиловая
кислота ()
Предполагают, что минорные нуклеотидные остатки защищают тРНК от атаки рибонуклеазами, принимают участие в кодировании аминокислот и важны для узнавания аминоацил-тРНК-синтетазой нужной тРНК.
В тРНК заметно преобладает сумма Г и Ц над суммой А и У. Молекулярные массы тРНК 17–35 тыс.
Первичную структуру тРНК впервые изучили американский биохимик Р. У. Холли (1965 г., тРНКала пекарских дрожжей) и отечественный биохимик А. А. Баев (1967 г., тРНКвал дрожжей). С тех пор определены первичные структуры более 100 различных тРНК. Это позволило выявить некоторые закономерности первичной структуры тРНК:
1. Все тРНК начинаются с фосфорилированного 5-конца; в 75% случаев первым основанием является гуанин.
2. На 3-конце всегда присутствуют три основания – ЦЦА, причем остаток А служит для связывания (акцептирования) аминокислоты.
3. В первичной структуре тРНК имеются гомологичные блоки, крайне близкие по чередованию нуклеотидных остатков. Кроме того, минорные нуклеотидные остатки закономерно распределены в первичной структуре тРНК.
Вторичная структура тРНК называется «клеверный лист»:
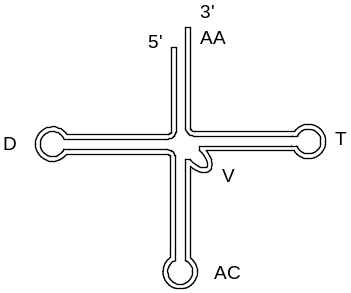
Эта структура характеризуется пятью ответвлениями («лепестками»), в составе которых имеется одноцепочечный участок и двухцепочечный стебель. Каждому ответвлению присвоено свое название либо по структурному, либо по функциональному признаку:
АА – аминоацильный (акцепторный) конец, к нему присоединяется аминокислота;
АС – антикодоновая петля, содержит антикодон;
Т – псевдоуридиловая петля, в этой петле содержится общая для всех тРНК последовательность ГТЦ; ответственна за связывание с рибосомой;
D – дигидроуридиловая петля, содержит дигидроуридиловую кислоту:
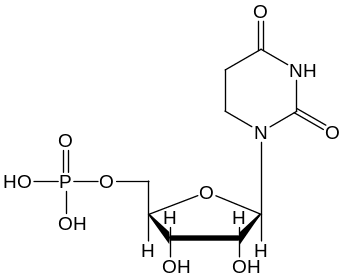
и АГ-последовательности;
V – вариабельная (добавочная) петля имеет для всех тРНК разные размеры или может отсутствовать.
Функции всех этих участков полностью не выяснены.
Третичная структура тРНК. Способ образования третичной структуры тРНК стал ясен после рентгеноструктурного анализа тРНКфен из дрожжей, впервые полученной в кристаллическом состоянии. Рентгеноструктурные исследования были выполнены одновременно в лабораториях американского ученого А. Рича и английского кристаллографа А. Клуга. Ее третичная структура напоминает букву L. Принципы образования третичной структуры тРНК, общие для всех классов РНК, обсуждались выше.
Рибосомальная РНК
У всех организмов найдено три вида рРНК, отличающиеся по молекулярным массам и локализации в рибосомах, обязательной составной частью которых они являются. Две рРНК высокомолекулярны, а третья сравнительно низкополимерна. Кроме того, в рибосомах эукариот присутствует еще одна низкомолекулярная рРНК.
Различают:
– 16–18S рРНК, локализованную в 30–40S субчастицах рибосом (Мr = 0,55–0,79106);
– 23–29S рРНК, локализованную в 50–60S субчастицах рибосом (Мr = 1,07–1,6106);
– 5S рРНК присутствует во всех рибосомах и локализована в 50–60S субчастицах (Мr = 40 тыс.);
– 5,8S рРНК характерна только для эукариотических рибосом (Мr ~ 50 тыс.).
Первичная структура рРНК. Нуклеотидный состав высокополимерных рРНК варьирует в довольно широких пределах и по мере усложнения организма все более смещается в сторону преобладания ГЦ-пар. рРНК митохондрий отличаются резким преобладанием АУ-пар (иногда их выделяют в отдельный класс – митохондриальная РНК). Высокомолекулярные рРНК содержат в 2–5 раз меньше минорных оснований, чем тРНК.
В 5S рРНК совершенно нет минорных оснований (иногда – только псевдоуридиловая кислота). 5S рРНК относится к ГЦ-типу. 5S рРНК содержит 120 нуклеотидных остатков (за редким исключением), последовательность которых в незначительной мере отличается у разных организмов.
Молекулы 5,8S рРНК в подавляющем большинстве случаев содержат 158 нуклеотидных остатков.
Работы по выяснению первичной структуры высокомолекулярных рРНК начаты в 1974 г. К настоящему времени раскрыты первичные структуры более сотни 16–18S рРНК и некоторых 23–29S рРНК.
Вторичная структура рРНК характеризуется спирализацией самой на себя полирибонуклеотидной цепи за счет взаимодействия комплементарных (Г–Ц и А–У) оснований, в результате в молекуле возникает различное число биспиральных участков (больше в 5S и 5,8S рРНК и меньше в 16–18S рРНК).
Третичная структура рРНК образуется за счет укладки плоскостных биспиральных и линейных участков молекул в более компактные структуры.
Функциональная роль рРНК: 16–18S и 23–29S рРНК являются структурной основой для формирования рибонуклеопротеинового тяжа, который, складываясь в пространстве, дает начало 30–40S и 50–60S субчастицам рибосом. Кроме того, они взаимодействуют с мРНК и аминоацил-тРНК; их участки распознаются белковыми факторами, принимающими участие в синтезе белка на рибосомах. Также они могут контактировать друг с другом при образовании 70–80S рибосом из 30–40S и 50–60S субчастиц и формировании активного центра рибосомы. 5S рРНК участвует в связывании тРНК с рибосомой за счет псевдоуридиловой петли.
Информационные РНК
Существование иРНК было предсказано в 1958 г. А. Н. Белозерским и А. С. Спириным. В 1962 г. мРНК была выделена в свободном виде.
Молекулярные массы мРНК варьируют от нескольких сотен тысяч до нескольких миллионов.
Функция иРНК заключается в копировании генетической информации с ДНК и переводе ее в аминокислотную последовательность (иРНК является матрицей для синтеза белка).
Одна молекула иРНК может содержать информацию об одном белке – моноцистронные иРНК, или кодировать биосинтез нескольких функционально взаимосвязанных белков – полицистронные иРНК.
Нуклеотидный состав иРНК крайне разнообразен.
Первичная структура иРНК. В составе иРНК есть информативные, работающие как матрицы в процессе биосинтеза белка, зоны и неинформативные участки:
– полиадениловые фрагменты длиной от 50 до 400 нуклеотидных остатков (кроме гистоновых иРНК) на 3-конце, которые участвуют в процессе созревания иРНК, предопределяют время жизни иРНК, способствуют переносу ее из ядра в цитоплазму и принимают участие в трансляции;
– небольшие повторяющиеся последовательности по 30 нуклеотидных остатков, располагающиеся неподалеку от полиаденилового фрагмента и необходимые для взаимодействия иРНК с рибосомой или отдельными белковыми факторами;
– кэп на 5-конце – нуклеотидная последовательность, азотистые основания в которой метилированы, а один из нуклеотидных остатков – 7-метилгуанозин, присоединен через трифосфатную группировку; кэп нужен для защиты иРНК от экзонуклеаз, для связывания белковых факторов при взаимодействии с рРНК. Он играет сигнальную роль при присоединении иРНК к рибосоме и участвует в трансляции.
Информативный фрагмент (транслируемый) начинается с АУГ или ГУГ на 5-конце и заканчивается УГА, УАГ или УАА на 3-конце.
АУГ или ГУГ соответствует антикодон формилметионил-тРНК; УАА, УГА и УАГ являются терминирующими трансляцию кодонами.
Вторичная структура иРНК полностью не выяснена, но известно, что полинуклеотидная цепь иРНК спирализована сама на себя. О третичной структуре иРНК известно только то, что она менее компактна, чем у рРНК.