

6.5.2. Вторичная структура днк
Молекулы природной ДНК в подавляющем большинстве случаев (кроме ДНК фагов) составлены парами взаимозакрученных полидезоксирибонуклеотидных цепей, каждой из которых свойственно специфическое, но противоположное чередование нуклеотидных остатков.
Модель структуры ДНК была впервые предложена в 1953 г. американским биохимиком Дж. Уотсоном и английским физиком Ф. Криком. Согласно этой модели, молекула ДНК состоит из 2-х полинуклеотидных цепей, которые образуют правую спираль (точнее – винтовую линию) относительно одной и той же оси; отсюда название – двойная спираль (рис. 15).
 Рис.
15. Двойная спираль ДНК
Рис.
15. Двойная спираль ДНК
Направление цепей взаимно противоположное (5–3 и 3–5). Сахарофосфатный остов располагается по периферии двойной спирали, а азотистые основания находятся внутри, и их плоскости перпендикулярны оси спирали.
Анализируя данные по нуклеотидному составу нуклеиновых кислот, американский биохимик Э. Чаргафф в 1950 г. сформулировал ряд правил.
1. У ДНК молярная сумма Г и А (пуриновые основания) равна молярной сумме Ц и Т (пиримидиновые основания). Эта закономерность несвойственна РНК.
2. В молекулах ДНК число остатков А всегда равно числу остатков Т. В таком же отношении находятся Г и Ц. В молекулах РНК этого нет.
Эти правила известны как принцип комплементарности или дополнения. Между азотистыми основаниями образуются специфические водородные связи, в результате чего осуществляется так называемое уотсон-криковское спаривание. А всегда образует водородные связи с Т, а Г – с Ц:

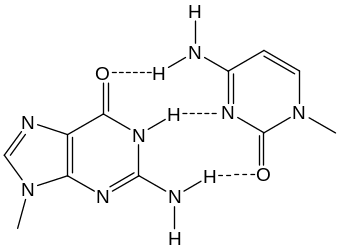
Аденин Тимин Гуанин Цитозин
Таким образом, более объемные пурины всегда спариваются с пиримидинами, имеющими меньшие размеры. Это приводит к тому, что расстояния между С1-атомами дезоксирибозы в двух цепях оказываются одинаковыми для АТ- и ЦГ-пар и равными 1,085 нм. В результате АТ- и ЦГ-пары включаются в двойную спираль без какого-либо заметного изменения геометрии остова. К тому же такое спаривание оказывается и более выгодным энергетически, т. к. реализуется максимальное число возможных водородных связей (2 между А и Т и 3 – между Г и Ц).
3. Отношение
суммы молярных концентраций Г и Ц к
сумме молярных концентраций А и Т у ДНК
и А и У у РНК у нуклеиновых кислот сильно
варьирует. Отношение
![]() для ДНК и
для ДНК и
![]() для РНК называется коэффициентом
специфичности нуклеиновых кислот.
для РНК называется коэффициентом
специфичности нуклеиновых кислот.
Геометрия двойной спирали. Соседние пары оснований находятся друг от друга на расстоянии 0,34 нм и повернуты на 36 вокруг оси спирали (рис. 16). На один виток спирали приходится, следовательно, 360/36 = 10 пар оснований, и шаг спирали равен 100,34 = 3,4 (нм). Диаметр двойной спирали равен примерно 20 нм. В двойной спирали ДНК образуются желобки (большой и малый). Это связано с тем, что сахарофосфатный остов расположен дальше от оси спирали, чем основания.
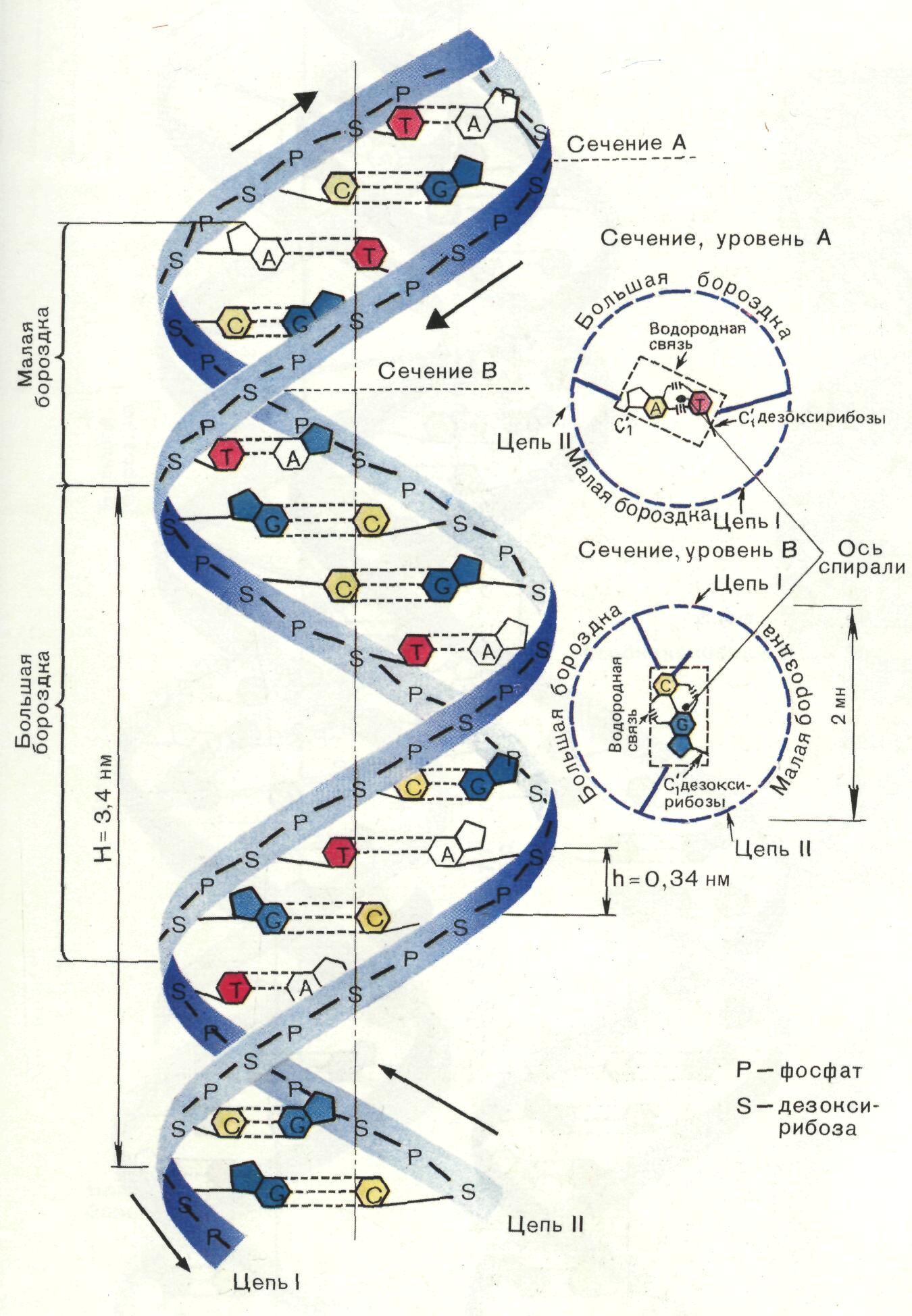 Рис. 16. Геометрия
двойной спирали ДНК
Рис. 16. Геометрия
двойной спирали ДНК
Стабильность двойной спирали обусловлена разными взаимодействиями. Отчасти за нее ответственны водородные связи между основаниями. Однако, по-видимому, более важную роль играет межплоскостное стэкинг-взаимодействие (от англ. stack – стог, куча, груда, стопка). Стэкинг-взаимодействия направлены вдоль оси спирали. При этом обеспечиваются не только выгодные ван-дер-ваальсовы контакты между атомами, но и возникает дополнительная стабилизация благодаря перекрыванию -орбиталей атомов контактирующих оснований. Стабилизация осуществляется также за счет благоприятного гидрофобного эффекта, проявляющегося в том, что неполярные основания защищены от непосредственного контакта с растворителем. Напротив, сахарофосфатный остов с его полярными группами и заряженными атомами экспонирован, что также стабилизирует структуру.
Биспиральные структуры в молекулах ДНК возникают не только при взаимодействии двух комплементарных цепей, но и в пределах одной и той же цепи. Это происходит в тех случаях, когда в комплементарных цепях ДНК присутствуют палиндромы (от греч. палин – обратно, дроме – бегу) – «обратно бегущие» последовательности нуклеотидных звеньев. Эти последовательности ДНК одинаково читаются как в прямом, так и в обратном направлении. Например:
ГАЦАЦЦАГЦТГГТГТЦ
ЦТГТГГТЦГАЦЦАЦАГ
Палиндромы характерны для тех участков молекулы ДНК, где расположены зоны «узнавания» структур ДНК ферментами и регуляторными белками. Эти участки могут спирализоваться сами на себя, образуя крестообразные структуры – шпильки.