

10.2.3. Конечные продукты распада аминокислот
В результате распада аминокислот возникают СО2, NН3, амины, кетокислоты и другие сложные органические вещества. Все они (кроме СО2 и NH3) подвергаются дальнейшей деструкции.
Амины путем окислительного дезаминирования превращаются в карбоновые кислоты:
R–CH2–NH2 + H2O + CO2 R–CHO + NH3 + H2O2 (фермент моноаминооксидаза)
R–CHO + H2O + НАД R–СООН + НАДН2 (фермент альдегиддегидрогеназа).
Аналогично идет реакция окислительного дезаминирования диаминов при посредстве диаминоксидаз.
Кетокислоты и карбоновые кислоты постепенно окисляются до СО2 и Н2О (см. обмен липидов).
Таким образом, конечными продуктами распада аминокислот являются Н2О, СО2 и NН3. Вода поступает в общий метаболический фонд, углекислый газ выводится из организма.
Аммиак является высокотоксичным соединением, поэтому концентрация его в организме должна сохраняться на низком уровне (1–2 мг/л в крови; концентрация 50 мг/л является летальной).
Поэтому аммиак должен подвергаться связыванию в тканях с образованием нетоксичных соединений, легко выделяемых с мочой.
Пути устранения аммиака в организме
Аммиак в организме может устраняться следующими путями:
1. Образование аммонийных солей: 2NH3 + H2O + CO2 (NH4)2CO3 (у некоторых обитателей гидросферы – пиявок, крабов, раков и др.).
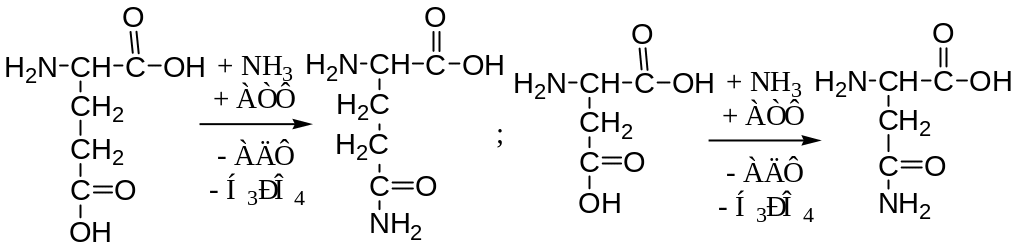
2. Биосинтез глутамина (и аспарагина). Это происходит в мозге, сетчатке, почках, печени и мышцах. Амиды выполняют транспортную функцию.
Синтез амидов требует энергии АТФ и присутствия глу или асп, свободного аммиака и катализируется глутамин- и аспарагинсинтетазами:
3. Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины. Образующаяся мочевина выводится с мочой, на долю ее приходится до 80–85% всего азота мочи. Синтез мочевины локализован в печени. Механизм синтеза мочевины – орнитиновый цикл мочевинообразования – описал Х. А. Кребс в 1932 г.
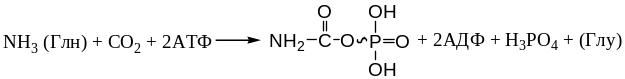
Образование мочевины начинается с синтеза макроэргического соединения карбамилфосфорной кислоты (карбамилфосфата) из СО2 и NH3 (или глн в качестве донора аммиака) (фермент карбамоилфосфатсинтаза):
Образовавшийся карбамилфосфат включается в специфический цикл, в котором в качестве регенерируемого компонента («катализатора») принимает участие аминокислота орн, которая не относится к числу аминокислот, формирующих белки, но является важным компонентом азотного обмена. Орн легко возникает при гидролизе арг.
Реакции цикла мочевины:
1. Конденсация карбамилфосфата и орн с образованием цитруллина. Фермент – орнитин-карбамилтрансфераза.
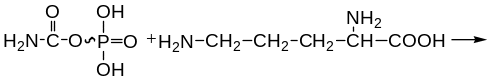
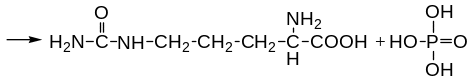
2. Взаимодействие цитруллина с асп с образованием аргининянтарной кислоты (аргининсукцината) Реакция сопряжена с гидролизом АТФ. Фермент – аргининсукцинатсинтетаза.
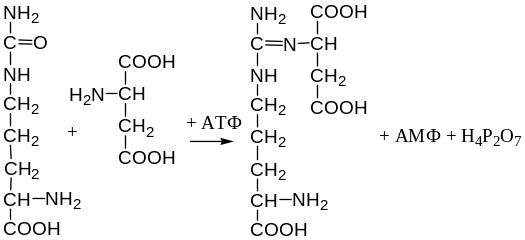
3. Отщепление фумаровой кислоты от аргининянтарной, катализируемое аргининсукцинатлиазой.
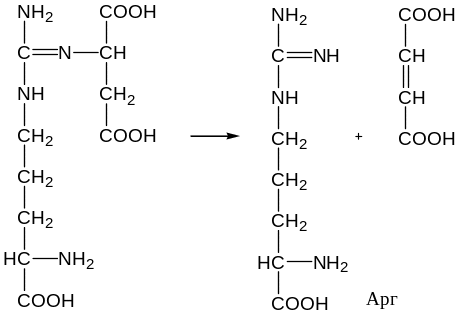
4. Гидролиз аргинина с помощью фермента аргиназы.
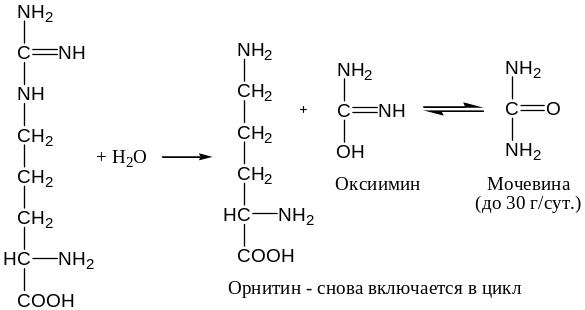
Суммарное уравнение синтеза мочевины:
СО2 + NH3 + 3АТФ + 2Н2О + асп
Н2N–CO–NH2 + 2АДФ + АМФ + фумарат + 2Н3РО4 + Н4Р2О7; -