

12.5. Взаимосвязь обмена углеводов и липидов
Связующие звенья – ПВК и ацетил-КоА.
1. Углеводы ПВК ацетил-КоА в.ж.к., стеролы и др.
фосфоглицерин липиды
Например:
ПВК
![]() 3-ФГА ⇄
ДОАФ
3-ФГА ⇄
ДОАФ
![]() 3-фосфоглицерин
3-фосфоглицерин
Фермент глицерол-3-фосфатдегидрогеназа
2. Липиды ацетил-КоА ПВК углеводы
глицерин 3-ФГА углеводы
У бактерий (фотосинтезирующих и аэробных):
ацетил-КоА ПВК + КоА
(Вместо НАД и НАДФ у них используется ферредоксин Fd. Ферредоксины – железосерные белки, содержащие негемовое железо: Fe2+ ⇄ Fe3+).
Помимо отмеченного выше, вещества, образующиеся в процессе обмена соединений одного класса, оказывают глубочайшее влияние на обмен веществ другого класса.
Пример: образование и-РНК, служащей матрицей для синтеза специфических белков, с одной стороны, и блокирование синтеза и-РНК определенного вида белками – с другой.
Таким образом, обмены белков, нуклеиновых кислот, углеводов, липидов и других соединений взаимозависимы, взаимообусловлены и составляют единое целое.
13. Регуляция обмена веществ
Общий ход биохимических процессов в организме регулируется внутренними и внешними факторами. Поэтому обмен веществ представляет собой единое неразрывное целое, и сам организм выглядит как самонастраивающаяся, саморегулирующаяся система, поддерживающая свое собственное существование путем обмена веществ.
Выделяют следующие уровни регуляции обмена веществ (в иерархическом порядке).
I. Метаболитный уровень регуляции
Слаженность обмена веществ во многом определяется концентрацией различных метаболитов или веществ, поступающих в организм с пищей.
Формы регуляции обмена веществ при участии метаболитов
1. Ускорение или замедление биохимических процессов за счет недостатка или избытка участников соответствующих реакций.
Например, объем белкового синтеза лимитируется поступлением в организм незаменимых аминокислот и интенсивностью синтеза заменимых.
2. Регуляция за счет конкурентных взаимоотношений обменных процессов, замыкающихся на общие (ключевые) метаболиты: ПВК, ЩУК, -кетоглутаровую кислоту, ацетил-КоА, Г-6-Ф.
Примеры:
– ПВК ⇄ лактат (отсутствие О2)
ацетил-КоА (наличие О2)
в восстановительных условиях,
при избытке НАДН2 и в присутствии ацил-КоА
– ПВК 3-ФГА ⇄ ДОАФ глицерофосфат фосфолипиды в окислительных условиях, при избытке НАД
3. Регуляция с помощью биологически активных веществ – витаминов, антивитаминов, коферментов, гормонов, антигормонов, вторичных посредников и др.
Метаболиты, взаимодействуя с ферментами, способны активировать или ингибировать их активность.
II. Оперонный уровень регуляции
Оперон – упорядоченная компактная совокупность цистронов (генов), считываемая как единое целое в процессе транскрипции иРНК на ДНК. Оперон может быть моноцистронным, если на нем синтезируется иРНК, предназначенная для синтеза одного белка, и полицистронным, если на нем синтезируется ряд иРНК.
На уровне оперона регулируется главным образом объем биосинтеза ферментов за счет изменения количества молекул иРНК, возникающих при транскрипции.
Возможны 2 пути регуляции биосинтеза ферментов на уровне оперона (подробно изучены у бактерий):
1. Индукция
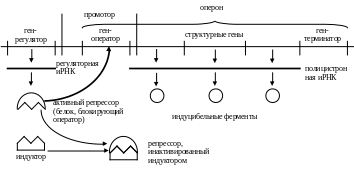
Клетки могут синтезировать специфические ферменты в ответ на присутствие специфических низкомолекулярных индукторов.
Индукторами могут быть:
– субстраты для индуцибельного(ых) фермента(ов) (лактоза для -галактозидазы у Escherichia coli);
– соединения, структурно сходные с субстратами индуцибельных ферментов.
Если одно соединение индуцирует сразу несколько ферментов данного катаболического пути (полицистронный оперон), то индукцию называют координированной.
Примеры индуцибельных ферментов у млекопитающих: тирозин--кетоглутарат-трансаминаза, ферменты цикла мочевины, цитохром Р-450 и некоторые другие.
Пример индукции. Клетки E. coli, выращенные на глюкозе, не способны сбраживать лактозу из-за отсутствия фермента -галактозидазы. Если в питательную среду добавить лактозу или другие -галактозиды, то индуцируется синтез -галактозидазы, и культура клеток обретает способность сбраживать лактозу.
– существуют специальные регуляторные белки, например, белок-активатор катаболизма (БАК, catabolite activating protein – CAP) у бактерий. БАК выступает в роли репрессора транскрипции сразу нескольких оперонов, общим для которых является то, что программируемые ими группы ферментов участвуют в первых стадиях использования нетрадиционных источников энергии – галактозы, арабинозы, лактозы. Непосредственным индуктором выступает в этом случае цАМФ. Если в клетке достаточно глюкозы, то цАМФ в значительных количествах не образуется, и все опероны оказываются в репрессированном состоянии. При истощении запасов глюкозы «включается» образование цАМФ и происходит индукция целого набора оперонов.
2. Репрессия
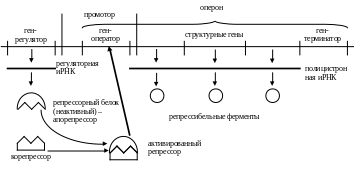
Клетки, способные синтезировать определенный метаболит, при наличии этого метаболита в среде могут приостановить его синтез в результате репрессии. В этом случае небольшая молекула (например, пурин, аминокислота) действует как корепрессор, блокируя синтез ферментов, участвующих в синтезе апорепрессора. После удаления из среды корепрессора, или же при истощении его запасов биосинтез соответствующих ферментов возобновляется – дерепрессия.
Корепрессором может быть:
– конечный продукт цепи реакций (репрессия по принципу обратной связи), например, гистидин для всех ферментов его биосинтеза у бактерий Salmonella typhimurium;
– промежуточное соединение в цепи (катаболитная репрессия).