

9.5. Общая схема распада углеводов
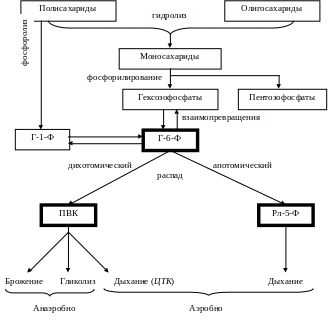
Г люкозо-6-фосфат
занимает центральное место в обмене
углеводов, а из дальнейших продуктов
узловые позиции принадлежат ПВК и
Рл-5-Ф.
люкозо-6-фосфат
занимает центральное место в обмене
углеводов, а из дальнейших продуктов
узловые позиции принадлежат ПВК и
Рл-5-Ф.
Зависимости между различными путями распада углеводов (брожение, гликолиз, дыхание; апотомический и дихотомический распад; анаэробный и аэробный) сложны и определяются как видовыми особенностями, так и условиями жизнедеятельности организмов.
Например, объем гликолиза в тканях находится в прямой зависимости от поступления кислорода: последний подавляет процесс образования молочной кислоты (эффект Пастера).
Даже в различных тканях и органах одного и того же организма соотношения путей распада углеводов могут быть различными.
Тем не менее можно установить некоторые общие закономерности:
У подавляющего большинства организмов аэробный путь распада углеводов в общем превалирует над анаэробным, а дыхание подавляет гликолиз и брожение.
Дихотомическому пути распада углеводов принадлежит в целом более видное место, чем апотомическому.
В значительной степени эти соотношения путей распада углеводов зависят от их энергетического эффекта.
9.6. Синтез углеводов
Синтез простых углеводов
Простые углеводы возникают главным образом при первичном биосинтезе органического вещества на Земле. Этот процесс осуществляется автотрофными организмами – растениями, фотосинтезирующими и хемосинтезирующими бактериями. Первичный синтез органического вещества в природе идет путем восстановления СО2 атмосферы с одновременным формированием органических молекул, новообразование которых сопровождается поглощением энергии.
В простейшем случае у хемосинтезирующих бактерий источником энергии служат реакции окисления неорганических соединений, например:
н
 итрозобактерии:
2NH3
+ 3O2
→ 2HNO2
+ 2H2O
итрозобактерии:
2NH3
+ 3O2
→ 2HNO2
+ 2H2Oнитробактерии: 2HNO2 + O2 → 2HNO3
серобактерии: 2S + 3O2 + 2H2O → 2H2SO4 (избыток кислорода)
2H2S + O2 → 2H2O + 2S (недостаток кислорода).
Фотосинтезирующие бактерии и зеленые растения используют для первичного синтеза органических веществ энергию световых лучей. Выделяют две фазы фотосинтеза:
I. Световая фаза: высвобождение атомов водорода:
НАДФ + Н2О
![]() НАДФН2
+ ½О2
НАДФН2
+ ½О2
и фосфорилирование АДФ (аналогично окислительному фосфорилированию):
АДФ + Н3РО4 АТФ.
II. Темновая фаза: суммарное уравнение:
6СО2
+ 12НАДФН2
![]() С6Н12О6
+ 12НАДФ + 12Н2О
С6Н12О6
+ 12НАДФ + 12Н2О
Или по стадиям:
Рл-5-Ф + АТФ Рл-1,5-дФ + АДФ (фермент фосфорибулокиназа)
Рл-1,5-дФ + СО2 2 3-ФГК (рибулозодифосфаткарбоксилаза)
2 3-ФГК + 2НАДФН2 2 3-ФГА + 2НАДФ + 2Н2О
3-ФГА ⇄ ДОАФ Ф-1,6-дФ Ф-6-Ф Г-6-Ф (обращение дихотомического пути).
Исходными веществами для синтеза углеводов гетеротрофными организмами могут служить продукты распада липидов, белков и других органических соединений.
Глюконеогенез – синтез глюкозы из неуглеводных продуктов. Предшественниками глюкозы в нем могут быть пировиноградная кислота или любое соединение, превращающееся в процессе катаболизма в нее или один из промежуточных продуктов цикла Кребса. У позвоночных наиболее интенсивно глюконеогенез протекает в печени и корковом веществе почек.
Большинство стадий глюконеогенеза представляет собой обращение реакций гликолиза. Только три реакции гликолиза (гексокиназная, фосфофруктокиназная и пируваткиназная) необратимы, поэтому в процессе глюконеогенеза на трех этапах используются другие ферменты:
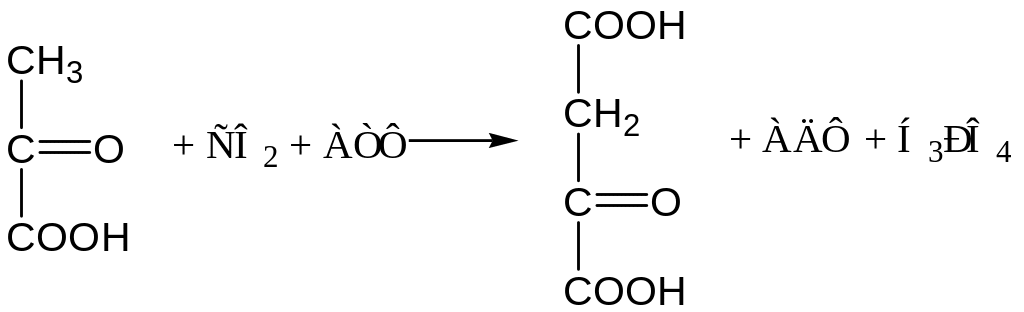
1. Образование фосфоенолпировиноградной кислоты из пировиноградной. Синтез ФЕП осуществляется в несколько этапов. Первоначально ПВК под влиянием пируваткарбоксилазы и при участии СО2 и АТФ карбоксилируется с образованием ЩУК:
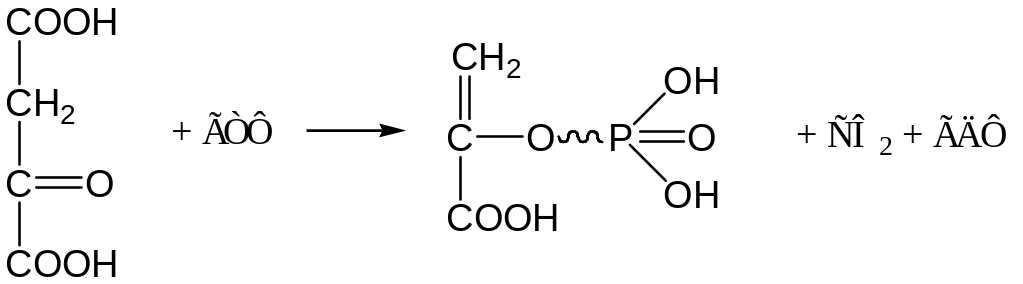
Затем ЩУК в результате декарбоксилирования и фосфорилирования под влиянием фосфоенолпируваткарбоксикиназы превращается в ФЕП. Донором остатка фосфорной кислоты служит ГТФ:
В процессе образования ФЕП участвуют ферменты цитоплазмы и митохондрий. Первый этап локализуется в митохондриях. Мембрана митохондрий непроницаема для образовавшейся ЩУК. Последняя здесь же восстанавливается до яблочной кислоты при участии НАД-зависимой малатдегидрогеназы (митохондриальной):
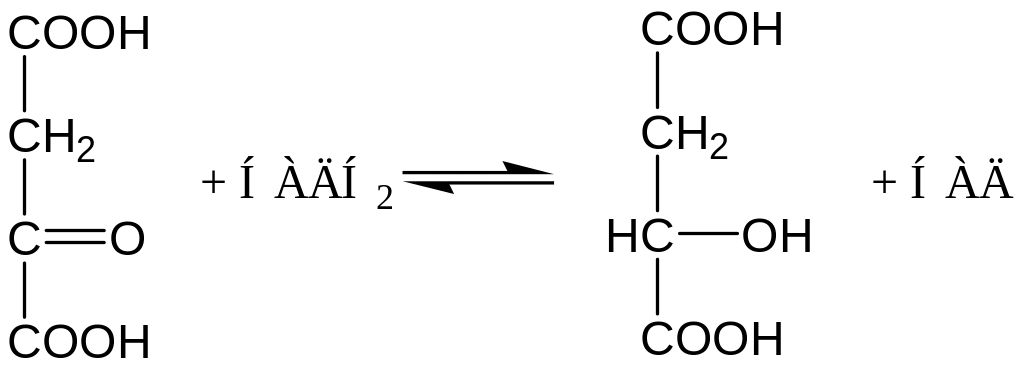
Яблочная кислота легко проходит через мембрану митохондрий в цитоплазму, где вновь окисляется при участии НАД-зависимой малатдегидрогеназы (цитоплазматической). Дальнейший процесс происходит в цитоплазме.
2. Превращение фруктозо-1,6-дифосфата во фруктозо-6-фосфат. ФЕП в результате ряда обратимых реакций гликолиза превращается во Ф-1,6-дФ. Далее глюконеогенез идет в обход необратимой эндэргонической реакции превращения Ф-1,6-дФ во Ф-6-Ф: Ф-1,6-дФ + Н2О Ф-6-Ф + Н3РО4.
Фермент фруктозодифосфатаза ингибируется АМФ и активируется АТФ (в отличие от фосфофруктокиназы, которая ингибируется АТФ и стимулируется АДФ и АМФ). Таким образом, когда концентрация АМФ мала, а концентрация АТФ велика, то стимулируется глюконеогенез. Напротив, когда величина отношения АТФ/АМФ низка, в клетке происходит расщепление глюкозы.
3. Образование глюкозы из Г-6-Ф. В последующей обратимой стадии биосинтеза глюкозы Ф-6-Ф превращается в Г-6-Ф, который может дефосфорилироваться (в обход гексокиназной реакции) под влиянием глюкозо-6-фосфатазы: Г-6-Ф + Н2О глюкоза + Н3РО4.
Между гликолизом, интенсивно протекающим в мышцах при их активной деятельности, и глюконеогенезом, особенно характерным для печени, существует тесная взаимосвязь. При максимальной активности мышц в результате усиления гликолиза образуется избыток молочной кислоты, диффундирующей в кровь. Значительная часть лактата в печени превращается в глюкозу (глюконеогенез). Образовавшаяся в печени глюкоза затем может быть использована как энергетический субстрат, необходимый для деятельности мышц.
Синтез олигосахаридов
Биосинтез олигосахаридов осуществляется путем реакций трансгликозилирования. Перенос гликозильного остатка на один моносахарид идет с фосфорного эфира другого моносахарида и ускоряется специфической гликозилтрансферазой.
Исходными соединениями, с которых в процессе синтеза олигосахаридов гликозильный остаток энергично переносится на моносахарид, служат нуклеозиддифосфатсахара, при гидролизе которых изменение уровня свободной энергии значительно выше, чем при гидролизе других доноров, а нуклеотидная часть их молекул способна обеспечить избирательность гликозилтрансферазной реакции.
Примеры. 1. Синтез сахарозы (идет только в растениях!) при участии фермента сахарозосинтазы:
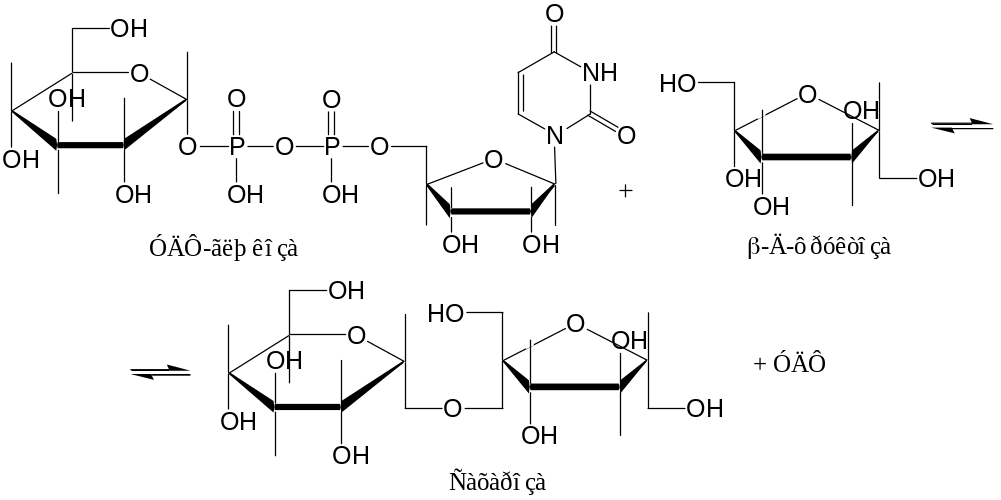
2. Синтез сахарозы при участии сахарозофосфатсинтазы:
УДФ-глюкоза + Ф-6-Ф УДФ + Сахарозо-6-Ф.
Далее действует фермент фосфатаза:
Сахарозо-6-Ф + Н2О → Сахароза + Н3РО4.
3. Синтез лактозы: УДФ-глюкоза + глюкоза → лактоза + УДФ.