

6.5.4.1. Репликация (редупликация, самоудвоение) днк
Репликация ДНК (от англ. replication – копирование; reduplication – удвоение) – это процесс, при котором информация, закодированная в последовательности нуклеотидов родительской ДНК, передается с максимальной точностью дочерней ДНК.
Биосинтез ДНК протекает из дезоксирибонуклеозидтрифосфатов при каталитическом воздействии ферментов ДНК-полимераз, которые катализируют перенос дезоксирибонуклеотидных фрагментов от дезоксирибонуклеозид-5-трифосфатов на группу –ОН 3-концевого фрагмента растущей цепи (ферменты класса трансфераз, подкласса нуклеотидилтрансфераз):


Таким образом, происходят 2 сопряженных процесса – гидролиз макроэргической связи (G << 0) и образование фосфодиэфирной связи (G > 0).
Рассмотрим механизм репликации (рис. 18).
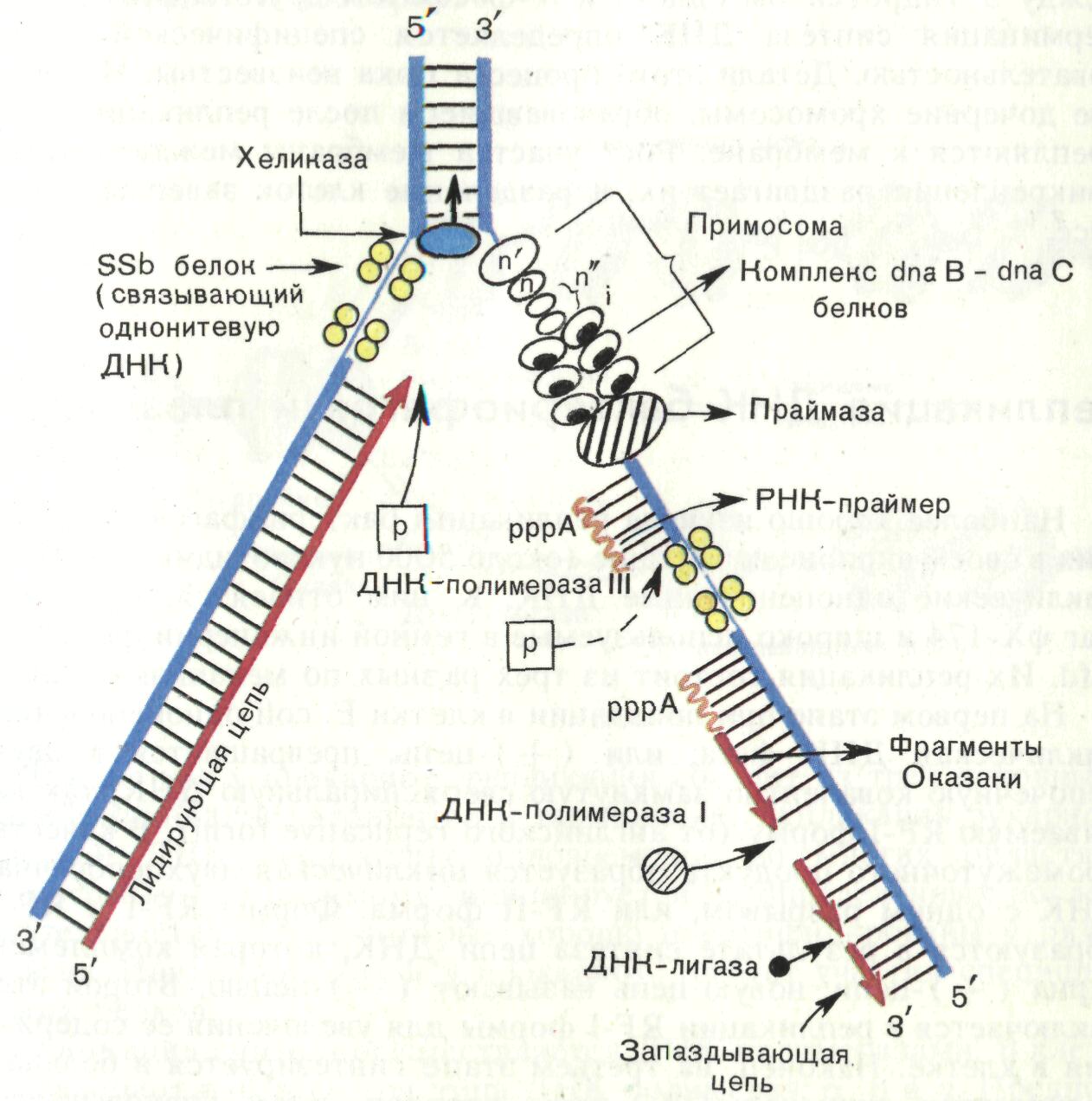
Рис. 18. Репликация ДНК (репликативная вилка)
1. Инициация синтеза (от англ. initiation – введение) – начало синтеза. Биосинтез ДНК начинается в строго определенном участке молекулы – участке ori (от англ. origin of replication) и от него распространяется в обе стороны.
Для инициации репликации ДНК расплетается в участке ori, образуя репликативную вилку. В этом участвуют ферменты геликазы, перемещающиеся вдоль молекулы ДНК в двух направлениях.
В отличие от геликаз, ДНК-полимеразы могут перемещаться только от 3- к 5-концу полинуклеотидной цепи, поэтому непрерывный синтез ДНК происходит только на одной нити ДНК – ведущей цепи.
На другой, запаздывающей (ведомой) цепи
1) должен образоваться затравочный нуклеотидный фрагмент – праймер (от англ. primer – запал), состоящий из рибонуклеотидов. В этом участвует фермент ДНК-зависимая РНК-полимераза (праймаза). Праймер образуется лишь в определенных участках запаздывающей цепи и состоит из нескольких десятков рибонуклеотидных звеньев.
2) Синтез ДНК идет в противоположном направлении (фермент – ДНК-полимераза).
2. Элонгация (от англ. elongate – удлинять) – продолжение синтеза.
На ведущей цепи происходит непрерывный синтез, а на запаздывающей – образуется новый праймер, т. е. синтез идет не непрерывно, а в виде фрагментов – фрагментов Т. Оказаки (1967 г.), состоящих из примерно 200 нуклеотидных остатков (соответствует длине нуклеосомных участков ДНК). Как только второй фрагмент Оказаки достигает первый, праймер I удаляется с помощью фермента РНКазы, а фрагменты «сшиваются» при посредстве фермента ДНК-лигазы.
Далее процессы повторяются.
3. Терминация (окончание) синтеза (от англ. termination – предел). Прекращение биосинтеза ДНК предположительно кодируется особой нуклеотидной последовательностью (палиндромом) на конце хромосомы. Кроме того, репликация прекращается при встрече двух репликативных вилок.
Закономерности репликации
1. Процесс репликации происходит по принципу комплементарности, чем достигается достаточно точная передача наследственной информации от родительской клетки к дочерней.
2. Молекула ДНК родительской клетки является матрицей, на которой синтезируются новые цепочки ДНК.
3. Как качественный, так и количественный нуклеотидный состав матричной и вновь синтезированной ДНК совпадают (точность – 1 ошибка на 1010 нуклеотидилтрансферазных реакций). Поэтому механизм репликации называют гомологическим.
4. Способ удвоения ДНК является полуконсервативным, так как в дочерних клетках одна цепь является родительской, а другая – вновь синтезированной:
