

10. Обмен белков
10.1. Переваривание белков
Главными источниками белков для человека являются пищевые продукты животного и растительного происхождения. Главным образом животные продукты (мясо, рыба, сыр) и только некоторые растительные продукты (горох, соя) богаты белками. Наиболее распространенные растительные пищевые продукты (хлеб, рис, макароны, гречневая крупа, пшено, орехи, картофель, капуста, морковь, свекла, яблоки, вишня и др.) содержат мало белка.
Белки пищи, за весьма редким исключением, не усваиваются организмом, если они не будут расщеплены в процессе переваривания пищи до свободных аминокислот, из которых затем в клетках организма строятся свойственные данному виду специфические белки.
Весь сложный процесс гидролиза пищевых белков, начиная от желудка и кончая тонким кишечником, «настроен» таким образом, чтобы путем последовательного действия протеолитических ферментов лишить белки пищи видовой специфичности и придать продуктам гидролиза способность всасываться в кровь через стенки кишечника (примерно 95–97% белков пищи всасываются в виде свободных аминокислот).
Таким образом, ферментативный аппарат желудочно-кишечного тракта осуществляет поэтапное, строго избирательное расщепление пептидных связей белковой молекулы:
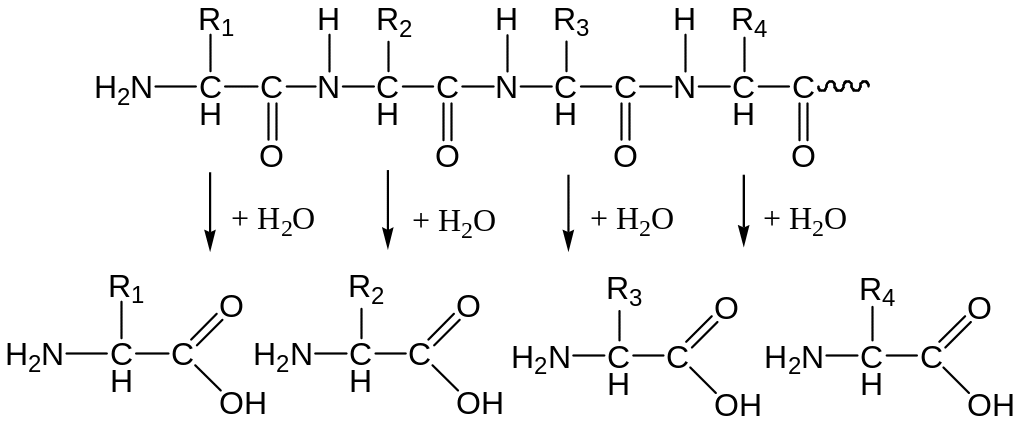
В желудке имеются все условия для переваривания белков. Во-первых, в желудочном соке содержится активный фермент пепсин. Во-вторых, благодаря наличию в желудочном соке соляной кислоты создается оптимальный для действия пепсина рН. Чистый желудочный сок имеет кислую среду (рН 0,9–1,6), а оптимум рН действия пепсина 1,5–2,5.
Соляная кислота переводит неактивный пепсиноген в активный пепсин, создает оптимальную среду для действия пепсина. В присутствии соляной кислоты происходят набухание белков (увеличение поверхности соприкосновения фермента с субстратом), частичная денатурация и, возможно, гидролиз пищевых белков.
Пепсин гидролизует преимущественно пептидные связи, образованные аминогруппами ароматических аминокислот (фен, тир). Он расщепляет практически все природные белки. В зависимости от природы белка, рН и скорости опорожнения желудка белки пищи в желудке подвергаются гидролизу пепсином с образованием полипептидов меньшего размера или коротких пептидов и, возможно, небольшого числа свободных аминокислот.
Дальнейшее превращение белков пищи осуществляется в тонком кишечнике, где на белки действуют ферменты панкреатического и кишечного соков. В поджелудочной железе вырабатываются трипсин, химотрипсин и карбоксипептидаза. Первые два фермента действуют на белки аналогично пепсину, но в отличие от него трипсин разрывает пептидные связи, образованные карбоксильными группами лиз и арг, а химотрипсин – карбоксильными группами ароматических аминокислот (фен, тир, три). Кроме того, они активны в слабощелочной среде (оптимум рН 7,2–7,8). Благодаря гидролитическому действию пепсина, трипсина и химотрипсина на белки образуются различной длины пептиды и некоторое количество свободных аминокислот.
Дальнейший гидролиз осуществляется под действием панкреатической карбоксипептидазы, кишечной аминопептидазы и различных дипептидаз. Точкой приложения аминопептидаз является пептидная связь с N-конца пептида. Карбоксипептидаза разрывает пептидную связь с С-конца пептида. Эти ферменты отщепляют по одной аминокислоте от полипептида, и, в конечном счете, остаются дипептиды, на которые действуют специфические дипептидазы, гидролизующие до свободных аминокислот.
Аминокислоты затем подвергаются всасыванию клетками кишечника.