

I.4. Группа гемма
Эта группа является окислительно-восстановительным кофактором ферментов дыхательной цепи, фотосинтеза и др.
Гем представляет собой простетическую группу. Он представляет собой порфириновый цикл, состоящий из 4-х пирольных колец, связанных группами СН, который в свою очередь связан ковалентными связями с белком. Ион Fe3+ , входящий в порфириновый цикл, способен восстанавливаться до иона Fe2+ и тем самым осуществлять перенос заряда, который является одним из важнейших процессов в живых клетках Электроны, переходят от одного вещества к другому иногда непосредственно, а иногда по цепи специальных переносчиков заряда. Принцип переноса заряда как раз и заключается в том, что ионы переходных металлов (Fe+2, Cu+2) в составе гема способны существовать в различных степенях окисления и принимать или отдавать электрны.
II. Коферменты переноса групп (трансферазы)
II.1. Аминотрансферазы.
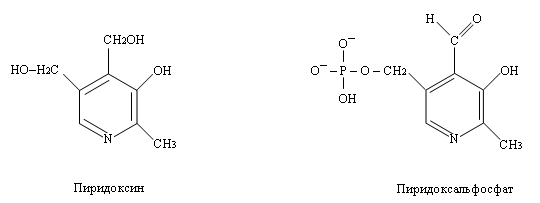
В метаболизме азотсодержащих соединений важную роль играют реакции переноса аминогрупп с остатков аминокислот на другие соединения (например, кетогруппы). Это реакции трансаминирования. Они катализируются ферментами, коферментом для которых служит пиридоксальфосфат (производная пиридоксина – витамина B6).
Пиридоксальфосфат ковалентно связан с ферментом за счёт взаимодействия альдегидной группы пиридоксальфосфата и аминогруппы аминокислоты лизина, входящей в состав белковой молекулы фермента (альдиминная связь).

Кроме этой связи пиридоксальфосфат связывается с белковой частью фермента за счёт ионных связей с участком фосфорного остатка и за счёт заряженного атома азота в пиридиновом кольце.
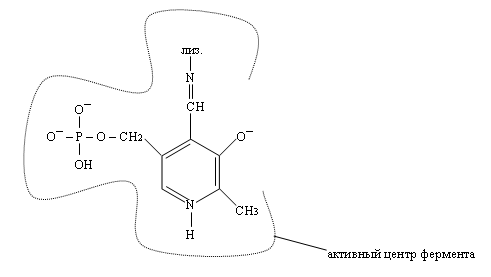
Соединение лизина и пиридоксальфосфата имеет плоское строение, обусловленное пиридиновым кольцом и дополнительной водородной связью между атомом N и атомом H гидроксильной группы.
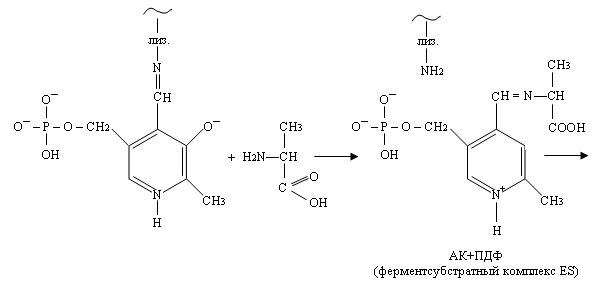
В процессе реакции трансаминирования аминокислоты, конкурируя с лизином, способны разрывать связь остатка лизина с карбонильной группой передоксальфосфата, вытесняя его из комплекса с ферментом.
Кофермент в этом случае играет роль посредника, т.е. промежуточного акцептора аминогруппы. Весть процесс переноса аминогруппы (NH2) с α-аминокислоты на α-кетокислоту происходит в несколько стадий:
1) На первой аминогруппа субстрата (например, аланина), конкурируя с лизином, образует альдиминную связь с передоксальфосфатом.
2) Внутримолекулярная перестройка. Под влиянием изомеразы двойная связь переходит от одного углерода к другому.

Образуется кетоиминная группировка – промежуточное соединение, у которого двойная связь находится у того атома углерода, который будет связан с кислородом на следующей стадии.
3) Стадия гидролиза. При этом образуется аминогруппа на коферменте и кетогруппа на бывшей аминокислоте.
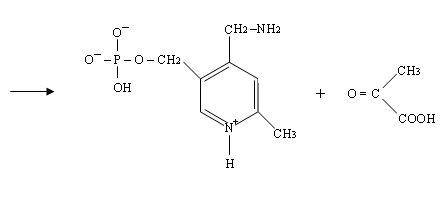
4) Четвёртая стадия происходит при наличии второго субстрата, которым является кетокислота, например, 2-оксоглутаровая:
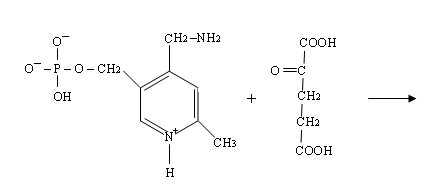
При этом происходит переход пиридоксальфосфата в исходную альдегидную группировку с лизином
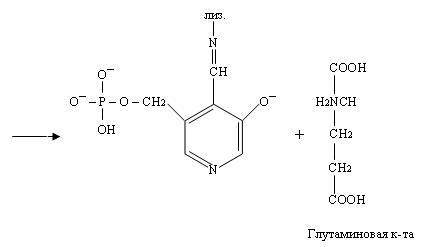
Как видно из уравнений, механизм реакции такой же, как при взаимодействии пиридина с аминокислотой, только конкурентами за кетогруппу кислоты становятся лизин и пиридоксальфосфат.
Осуществление рассмотренной последовательности реакций становится возможным благодаря возникающим конкурентным отношениям. По конкурирующим способностям за карбонильной группой пиридоксальфосфата аминокислоты располагаются в следующий ряд:
глутамат < остаток лизина в полипептиде < остаток аминокислоты
Суммарно весь этот процесс можно изобразить следующим образом:
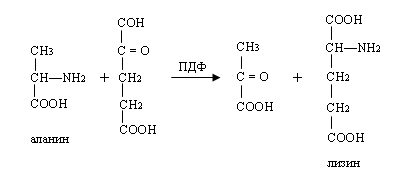
ПДФ в комплексе с декарбоксилазой может катализировать декарбоксилированные глутаминовые кислоты (образуется γ-аминомасляная кислота), гистодина, триптофана и др. аминокислоты.