

Онтогенетическое изменение темнового дыхания
На современном уровне знаний дыхание у растений рассматривается как механизм мобилизации запасенной энергии для функционирования биологических систем синтеза новых структур.
В целом организме существует очень много направлений, на которые может быть распределена энергия, получаемая растением в процессе дыхания Эти направления возможно представить следующей схемой (Рис.9.).

Рис.9. Трата энергии, образованной в результате процесса дыхания, на различные «нужды» растения»
Mc Cree (1970) на основе полученных им экспериментальных данных описал дыхание в виде двух составляющих - дыхание роста и дыхание поддержания (Рис.10).
Рис.10. Дыхание роста и дыхание поддержания
Дыхание роста обеспечивает энергией ростовую активность системы, а дыхание поддержания — всю ранее созданную биомассу. По сути, дыхание поддержания определяет скорость обновления структурной биомассы. Около 60% дыхания поддержания обусловлено обновлением белков (Головко,1999). В неблагоприятных условиях основные траты энергии приходятся на дыхание поддержания. У молодых растущих листьев эффективно дыхание роста, а дыхание поддержания невелико. Существует обратная пропорциональная зависимость между скоростью роста и дыханием поддержания: чем выше скорость роста, тем ниже дыхание поддержания.
У различных культур отношение дыхания к фотосинтезу в первую половину вегетации отличается мало. Однако, во второй половине вегетации, когда увеличивается доля дыхания от фотосинтеза — различия между разными культурами становятся существенными.
Итак:
-
наибольшей дыхательной активностью обладают ткани, находящиеся в состоянии активного деления,
-
органы с низким уровнем жизнедеятельности имеют слабую дыхательную активность.
Важным является период цветения и завязывания плодов, когда дыхание растений активизируется. Это связано с повышением доли активно делящихся тканей. В период цветения активизируется дыхательный газообмен всего растения, что обусловлено изменениями в обмене веществ всего организма при переходе к более поздним фазам развития. Увеличение дыхания во вторую половину вегетации происходит, в основном, за счет увеличения дыхания поддержания.
Принято считать, что, в среднем за вегетацию, величина темнового дыхания составляет примерно 20-50% от величины фотосинтеза. В онтогенезе прослеживается четкая связь между увеличением листовой поверхности растений и повышением его дыхания. С возрастом, в связи с образованием большой массы нефотосинтезирующих органов и уменьшением активности фотосинтетического аппарата, отношение выделяемого количества СО2 за ночь, к поглощенному за световой период, возрастает, достигая 52-61%.
Есть еще одно важное направление траты дыхательного субстрата — дыхание корней. Дыхание корней в течение вегетации может достигать 30% от дыхания всей биомассы, однако к концу вегетации оно снижается до 5%.
Суточная динамика интенсивности фотосинтеза
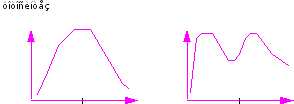
Наиболее интересным вопросом при изучении фотосинтетического газообмена является суточная динамика этого процесса. Впервые оценка выделения и поглощения СО2 в течение дня была сделана Mac Lean (1920). Экспериментируя на листьях кокосовой пальмы он обнаружил, что кривая поглощения углекислого газа имеет провал в полуденные часы. Наличие дневной ритмики фотосинтеза подтвердилось в дальнейшем работами других исследователей: Алексеева (1922-1924; Вотчала и Толмачева (1926), которые объясняли меняющееся в течение дня поглощение углекислого газа влиянием внешних факторов. Широкое изучение дневной ритмики фотосинтеза в разных климатических зонах (с учетом влияния разных факторов среды) было предпринято Костычевым с сотрудниками (1930). Этими авторами было установлено, что суточный ход фотосинтеза может быть выражен только двумя типами кривых: либо с одним максимумом в середине дня, либо с двумя максимумами — в утренние и послеполуденные часы (Рис. 11).
Однако, оказалось, что полученные кривые суточного хода фотосинтеза не повторяли суточного хода кривых изменения внешних факторов. Единственным выводом здесь могло быть существенное влияние на фотосинтез внутренних факторов, хотя, вполне вероятно, что динамика изменения внутренних факторов могла определяться изменением внешних факторов.
12 ч. 12 ч.
Один максимум два максимума
Рис. 11. Суточная динамика фотосинтеза
Работы по изучению суточной динамики фотосинтеза продолжались и в дальнейшем. К известным уже закономерностям добавилось представление о том, что возможно отсутствие видимой ритмики поглощения СО2 вообще в течение светлого периода суток. Многочисленные исследования в период с 1930 до 1970 гг. позволили собрать большой фактический материал и обобщить данные о наиболее общих закономерностях этого процесса.
Итак, было установлено, что в случае с одновершинной кривой фотосинтеза, максимум его интенсивности мог наблюдаться в 7-10 часов утра с дальнейшим постепенным спадом, либо около 12 часов.
В случае с двухвершинной динамикой наиболее интенсивный фотосинтез наблюдался в 6-10 часов утра, далее, в период наиболее интенсивной солнечной инсоляции - в 12 часов дня, наблюдалась депрессия фотосинтеза, после этого, к 15-16 (или к 17-19) часам интенсивность фотосинтеза вновь возрастала.
На современном уровне знаний принято считать, что динамика суточного хода фотосинтеза определяется комплексом внешних (экзогенных) и внутренних (эндогенных) факторов.
Логично связывать динамику интенсивности фотосинтеза с солнечным излучением, но именно тогда, когда солнечное излучение максимально, часто наблюдается полуденная депрессия фотосинтеза. Существовало несколько гипотез относительно того, чем это вызвано. В частности, это могло определяться недостатком воды в полуденные часы и активным закрыванием устьиц. Для подтверждения этой гипотезы были проведены эксперименты (Чмора, 1967) в условиях стабильной температуры и влажности. Оказалось, что при стабильной влажности у одних и тех же культур суточный ход фотосинтеза выражен кривой с одним максимумом в 10-11 часов, а в условиях недостаточного увлажнения — с двумя максимумами и с полуденной депрессией.
Казалось бы гипотеза доказана, но она не могла объяснить фактов полуденной депрессии у растений, лишенных устьиц, либо наличия двухвершинной кривой поглощения СО2 при достаточном водообеспечении (что иногда наблюдалось).
Ниловской с соавторами (1972) было проведено тщательное изучение суточной ритмики фотосинтеза в зависимости от тех эндогенных факторов, которые определяли внутренний режим растений, то есть от выработки определенного количества ассимилятов. В этих экспериментах прослеживается динамика изменения дневного хода фотосинтеза при изменении облученности растений.
Ниже приведены данные суточного газообмена посева моркови при 18-ти часовом облучении (Рис. 12).
А Б
6 9 12 15 18 21 24 6 9 12 15 18 21 24
более молодое растение более зрелое растение
(60 дн) (90 дн)
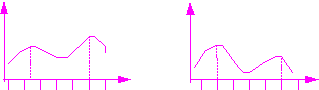 Рис.12.
Динамика изменения суточной ритмики
фотосинтеза у молодых (А) и развитых (Б)
растений моркови (Ниловская, 1972)
Рис.12.
Динамика изменения суточной ритмики
фотосинтеза у молодых (А) и развитых (Б)
растений моркови (Ниловская, 1972)
Фотосинтетическое поглощение растениями моркови углекислого газа имело вид двухвершинной кривой с двумя максимумами в 10 и 21 час. Изменение длительности светового периода до 18 и 24 часов позволило сделать следующие заключения:
-
При 18 часовом фотопериоде процесс фотосинтеза имеет колебательный характер. Для него характерна двухвершинная кривая.
-
При 24 часовом фотопериоде колебательный характер сохранялся, но кривая имела два максимума.
-
В ранний период вегетации растения откликаются на повышение облученности активнее.
В экспериментах, проведенных на различных культурах при разных режимах облучения, были установлены следующие закономерности:
-
существуют культуры, отрицательно реагирующие на непрерывное облучение в течение суток (к числу таких культур относятся томаты),
-
разные культуры имеют различную динамику хода фотосинтеза при непрерывном облучении- от одновершинной до много вершинной кривой,
-
регулирующее действие на ритмы фотосинтеза оказывает чередование светового и темнового периодов,
-
увеличение освещенности усугубляет величину депрессии фотосинтеза.
Последнее утверждение является очень важным для понимания природы колебательного характера дневного хода фотосинтеза. Активизация фотосинтеза в ответ на улучшение условий фотосинтетической деятельности обычно всегда сопровождается усилением депрессии фотосинтеза. Из этого можно сделать вывод о том, что внутри растения накапливаются неиспользованные метаболиты, тормозящие активность фотосинтеза. И чем активнее фотосинтез, тем интенсивнее их выработка. Действительно, существует так называемая система обратной связи, по которой активность работы фотосинтетического аппарата ограничивается избыточным накоплением углеводов.
Сама идея ограничения фотосинтеза ассимилятами очень стара. Она рассматривалась еще в 18 веке. Ее подтверждают эксперименты по введению в лист экзогенных (не произведенных самим листом) сахаров. При этом, после подобного введения наблюдается спад активности фотосинтеза. Считается, что саморегуляция фотосинтеза находится под четким контролем уровня метаболитов (Мокроносов,1970). Исключением являются случаи, когда возможен свободный отток ассимилятов в органы, их запасающие (временно депонирующие). Органами, способными к депонированию ассимилятов, являются корни, плоды, стебли, черешки листьев.
В настоящее время зависимость полуденной депрессии фотосинтеза от количества ассимилятов считается доказанной. Полагают, что при интенсивном освещении и, следовательно, интенсивном фотосинтезе величина полуденной депрессии будет значительной (Рис. 13). Если же освещенность понижена, то и ритмика фотосинтеза выражена слабо.
Возникает вопрос нужна ли растениям, испытывающим дневную депрессию фотосинтеза, в этот период высокая облученность, или (поскольку у растений снижается фотосинтез) в это время выгоднее поддерживать низкую облученность, не только для того, чтобы сэкономить на энергии, но и для того, чтобы помочь растениям использовать накопленные ассимиляты.
Рис.13. Величина депрессии фотосинтеза при благоприятных для его протекания условиях (А) и при условиях, ограничивающих его интенсивность (Б).
Интересные эксперименты, в которых изучалось влияние различных вариантов облучения были проведены на томатах (Боковая, 1971). В этих экспериментах оценивали влияние следующих режимов облученности:
1)130 Вт/м2 в течение всего светового периода
2)175-50-175 Вт/м2 со снижением облученности в период полуденной депрессии фотосинтеза
3)175 Вт/м2 в течение всего светового периода (Рис.14).
Поглощение СО2 (мг/час .дм-2)
А Б
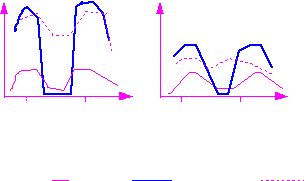
8час 20 час 9час 18 час
Рис 14. Изменение суточного ритма фотосинтеза у растений томата в возрасте 60 дн (А) и 90 дн (Б) при трех режимах облученности:
130 Вт/м2 175-50-175 Вт/м2 175 Вт/м2
В экспериментах было показано, что уменьшение облученности в период дневной депрессии резко усугубляло ее глубину, но не затягивало ее длительность. В этих опытах была подтверждена связь интенсивности фотосинтеза с регулирующим влиянием углеводов.
В результате было установлено, что интенсивность поглощения СО2 была несколько выше при чередующемся режиме облучения. Однако, анализ продуктивности показал, что томаты с постоянным облучением были более продуктивными. По-видимому, здесь играют роль еще какие-то факторы. Возможным фактором может быть интенсивность дыхания, которая активизировалась при чередовании облучения. Причина активизации дыхания могла быть вызвана либо зависимостью дыхания от количества углеводов, производимых при активации фотосинтеза, либо неблагоприятными условиями, складывающимися в момент снижения облученности, которое могло вызывать активацию дыхания, либо негативным влиянием смены облученности на дыхательные ферменты.
Ключевые слова:
онтогенез, ассимиляция, диссимиляция, дыхательный коэффициент,, дыхание роста, дыхание поддержания, полуденная депрессия фотосинтеза.