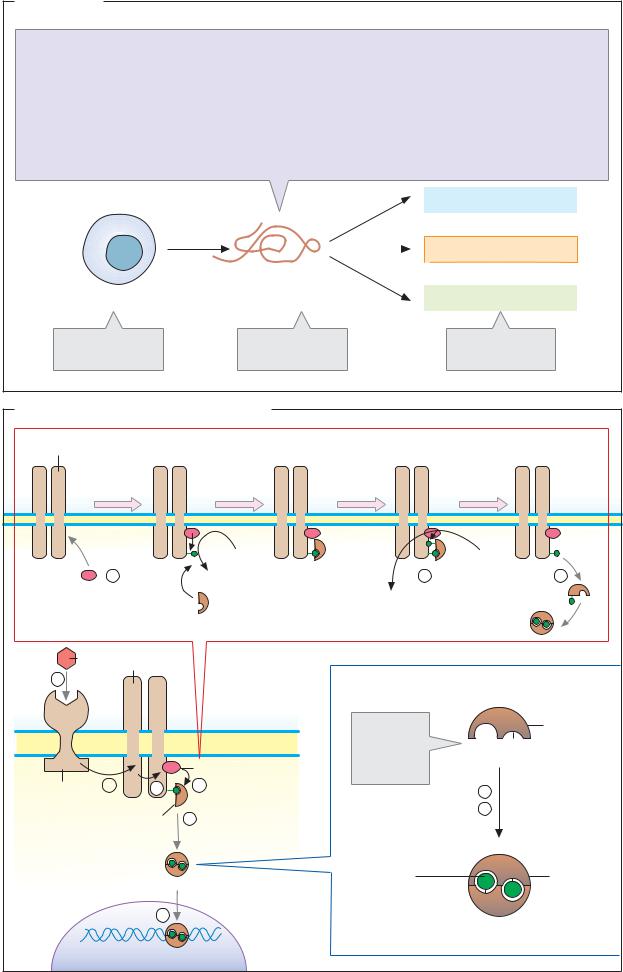

 permeability
permeability 


 Blood pressure
Blood pressure
 Blood glucose
Blood glucose  Lipolysis
Lipolysis
 Blood glucose
Blood glucose 

 ACTH
ACTH

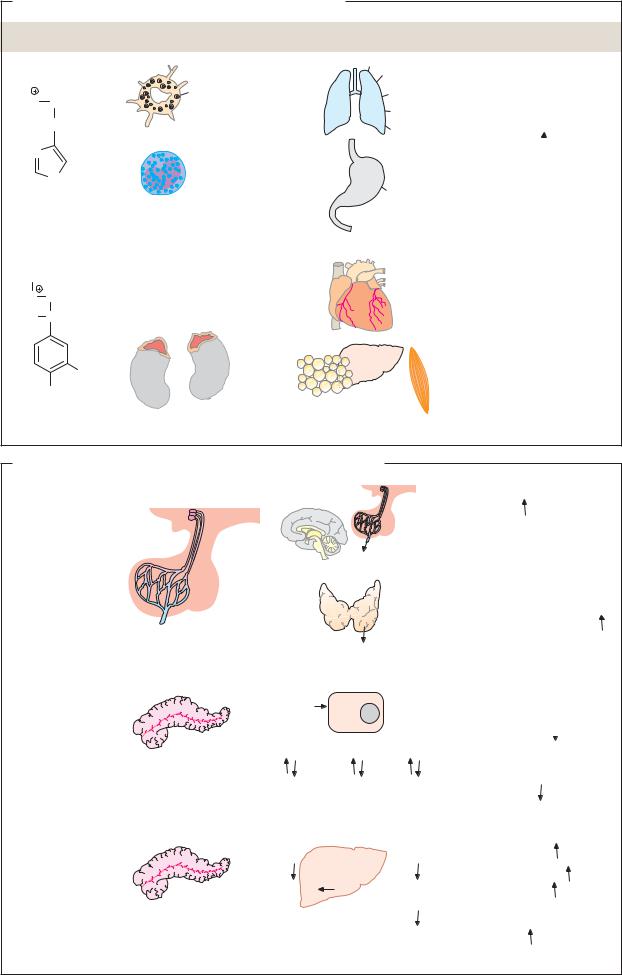
|
|
|
|
Hydrophilic hormones |
|
387 |
|||||
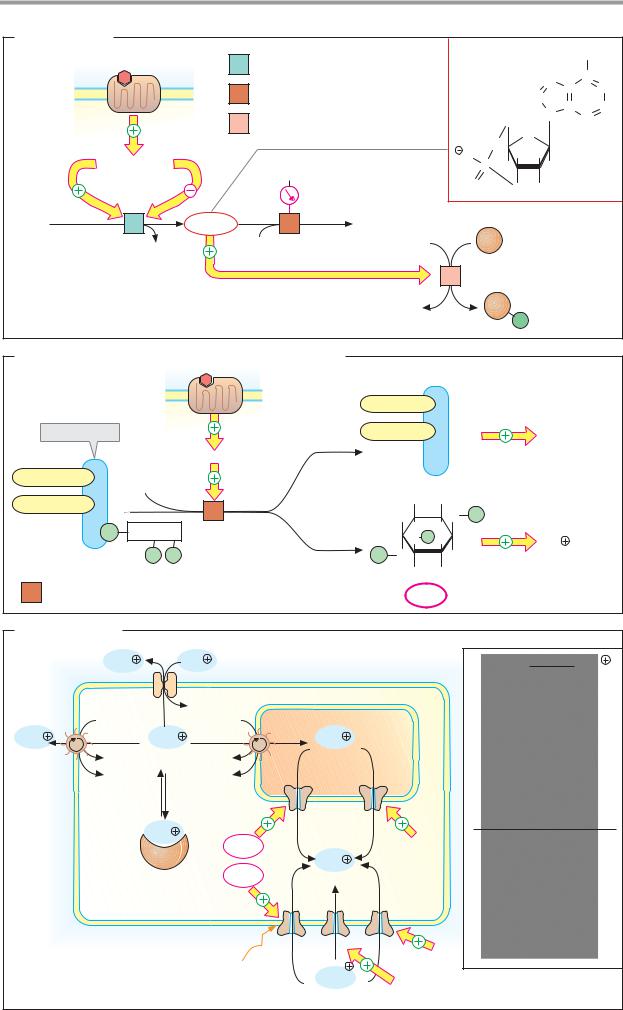
A. Cyclic AMP |
|
|
|
|
|
|
|
|
|
|
|
|
7 Helix receptors |
1 |
Adenylate cyclase 4.6.1.1 |
cAMP |
|
|
NH2 |
||||
|
|
|
C |
|
|||||||
|
|
|
|
|
|
|
|
|
N |
N |
|
|
|
2 |
Phosphodiesterase 3.1.4.17 |
|
|
|
|
C |
|||
|
|
|
|
|
HC |
C |
CH |
||||
|
|
3 Protein kinase A 2.7.1.37 |
|
|
5' |
N |
|||||
|
|
|
|
|
CH2 |
|
N |
|
|||
|
|
|
|
|
|
|
O |
O |
|
|
|
|
|
|
|
|
O |
|
H |
H |
|
|
|
|
G proteins |
|
|
|
P |
|
|
|
|||
|
|
Caffeine |
|
|
|
H 3' |
H |
|
|
||
|
Gs |
Gi |
|
O |
|
|
|
|
|||
|
|
|
|
|
O |
OH |
|
|
|||
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
||
ATP |
1 |
cAMP |
2 |
AMP |
|
|
|
Enzymes |
|
|
|
|
|
|
Transcription |
|
|||||||
|
PPi |
|
H2O |
ATP |
|
|
|
factors |
|
|
|
|
|
|
|
|
|
Ion channels |
|
||||
|
|
|
|
3 |
Protein kinase A |
|
|
||||
|
|
|
|
ADP |
|
|
|
P |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
B. Inositol 1,4,5-trisphosphate and diacylglycerol |
|
|
|
|||||||
|
|
7 Helix |
|
|
|
|
|
DAG (Diacylglycerol) |
||
|
|
receptors |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|||
|
Phospholipid |
|
|
|
|
|
|
|
Protein |
|
|
|
|
|
|
G protein (Gq) |
|
|
|
kinase C |
|
|
|
|
|
|
|
|
|
|
||
Acyl residue 1 |
Glycerol |
H2O |
|
|
|
|
|
|
||
|
|
|
|
|
OH |
OH |
|
|||
Acyl residue 2 |
|
|
|
4 |
|
|
||||
|
|
|
|
H |
O |
P |
||||
|
|
|
|
|
|
|
|
|||
|
|
P |
|
Inositol |
|
|
H |
H |
Intracellular |
|
|
|
|
|
|
O P |
H |
||||
|
|
|
|
P |
P |
|
|
Ca2 |
||
|
|
|
|
|
P |
O |
H |
release |
||
|
|
PlnsP2 |
|
|
|
H |
OH |
|
||
|
|
|
|
|
|
|
|
|
||
4 |
Phospholipase C 3.1.4.3 |
|
|
InsP3 |
|
|||||
C. Calcium ions |
|
|
|
|
|
|
|
|
||
|
|
Ca2 |
|
Na |
|
|
|
|
Ca2 |
|
|
|
ATP |
|
|
|
ATP |
ER/SR |
|
|
|
Ca2 |
|
|
|
Ca2 |
|
Ca2 |
|
|
|
|
|
|
ADP |
10-100 nM ADP |
|
|
|
|
|||
|
|
Pi |
|
|
|
Pi |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
a |
|
|
|
|
Ca2 |
|
InsP3 |
Ryanodine |
|
|
|
|
|
|
|
|
|
cAMP |
Ca2 |
|
|
|
|
|
|
|
Calcium- |
500-1000 nM |
|
|
|
||
|
|
|
|
binding |
|
|
|
|
|
|
|
|
|
|
protein |
|
|
|
|
|
|
|
|
|
|
|
Depolarization |
|
|
|
b |
|
|
|
|
|
|
|
|
Ca2 |
Glutamate |
|
|
1. Calcium transport |
|
|
ATP |
|
2. Calmodulin |
|||||
|
|
ca. 2 500 000 nM |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|

|
|
|
|
|
|
|
|
|
|
|
Hydrophilic hormones |
389 |
|||
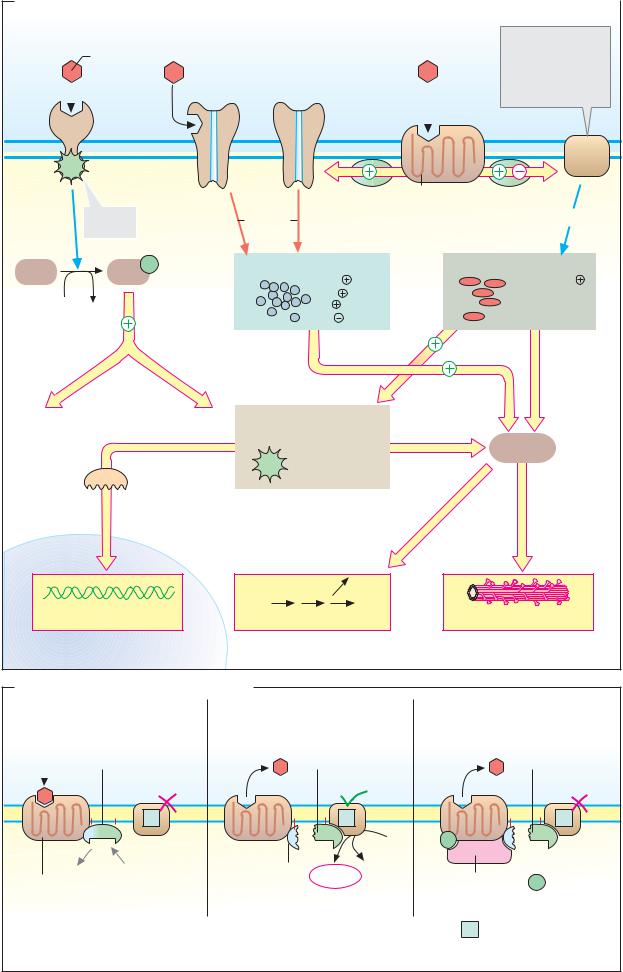
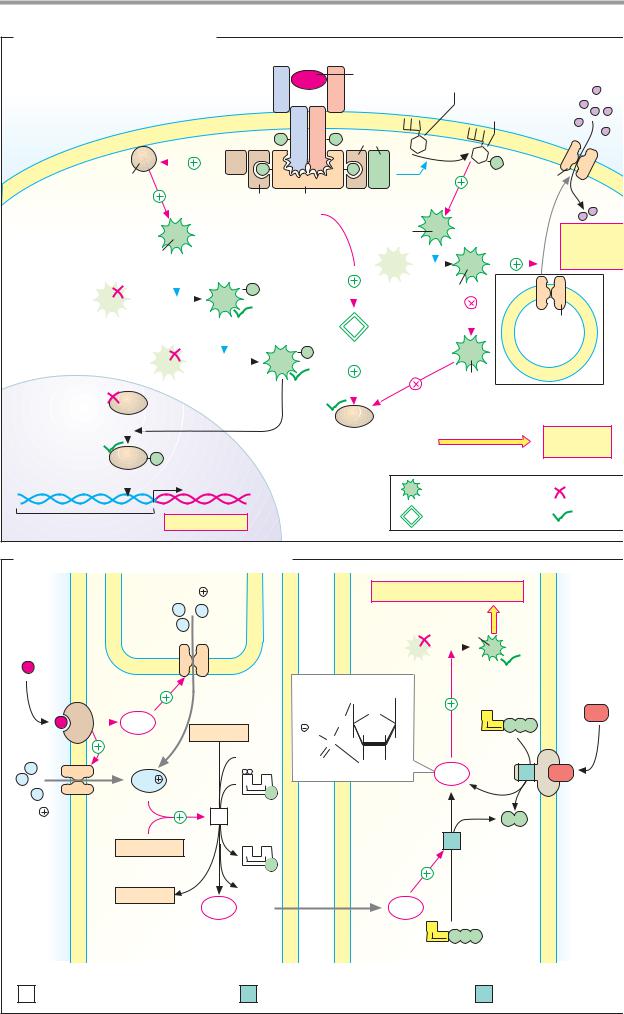
A. Insulin: signal transduction |
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
Insulin |
|
|
|
|
|
Insulin |
Phosphatidyl- |
|
|
Glucose |
||
|
|
|
receptor |
α |
|
α |
|
|
|
inositols |
|
Phosphatidyl- |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
inositol |
|
|
|
|
|
|
|
|
β |
β |
PI-3- |
|
|
3-phosphate |
|
|||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
SOS |
|
P |
|
P |
Kinase |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
P |
|
|
|
P |
|
|
|
|
P |
Glut-4 |
|
|
|
Ras |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
Grb-2 |
|
IRS-1 |
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
? |
|
|
PDK-1 |
|
|
|
Glucose |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
Raf |
|
|
|
|
|
|
|
|
|
|
|
|
uptake |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
MEK |
|
P |
|
|
|
|
|
|
|
PK-B |
|
|
|
|
|
(MAPKK) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
PP-1 |
|
|
|
Glut-4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
MAPK |
|
|
|
|
P |
|
|
|
|
|
|
|
|
|
|
(ERK) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
GSK-3 |
Intracellular |
|||
Transcription |
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
vesicle |
|
|||
factors |
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
Glycogen synthase |
|
|
Glycogen |
|||||
Activated |
P |
|
|
|
|
(active) |
|
|
|
|
synthesis |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
transcription |
|
DNA |
|
|
|
|
|
|
|
|
|
|
|
|
|
factors |
|
|
|
|
|
|
|
|
Protein kinase |
|
Inactive |
||||
|
|
|
|
|
|
Nucleus |
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
Protein phosphatase |
Active |
|||||
|
Promoter |
Transcription |
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
B. Nitrogen monoxide (NO) as a mediator |
|
|
|
|
|
|
|
|
|
||||||
|
|
Ca2 |
|
|
|
|
|
|
|
Physiological effects |
|
|
|||
Signaling |
ER |
|
|
|
|
|
|
|
|
PK-G |
|
|
|
|
|
substance |
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
5' |
Guanine |
|
|
|
ANP |
|||
|
|
|
|
|
|
|
|
CH2 |
|
|
|
|
|
|
|
|
|
InsP3 |
|
|
|
|
O |
|
O |
|
G |
|
|
|
|
|
|
Arginine |
|
|
|
|
H |
H |
|
|
P |
P P |
|
||
|
|
|
|
|
|
O |
|
|
GTP |
|
|
||||
|
|
|
2 O2 |
|
|
P |
H 3' |
H |
|
|
|
||||
|
|
|
|
|
O |
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
O |
OH |
cGMP |
|
|
3 |
|
|
|
|
Ca2 |
|
|
|
|
|
|
|
|
|
||||
|
|
N |
A |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
P |
|
|
|
|
|
|
|
|
|
|
|
Ca2 |
|
|
3/2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
P |
P |
|
|
|
|
Calmodulin |
3/2 |
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
N |
A |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
P |
|
|
|
|
|
|
|
|
|
|
|
|
|
Citrulline |
2 H2O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
NO· |
|
|
|
|
|
|
NO· |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
G |
GTP |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
P P P |
|
|
||
|
|
Endothelial cell |
|
|
|
|
|
|
|
Vascular muscle cell |
|
|
|||
1 |
NO synthase 1.14.13.39 |
2 |
Guanylate cyclase 4.6.1.2 |
|
3 |
ANF receptor 4.6.1.2 |
|||||||||

|
|
|
|
|
Other signaling substances |
391 |
|
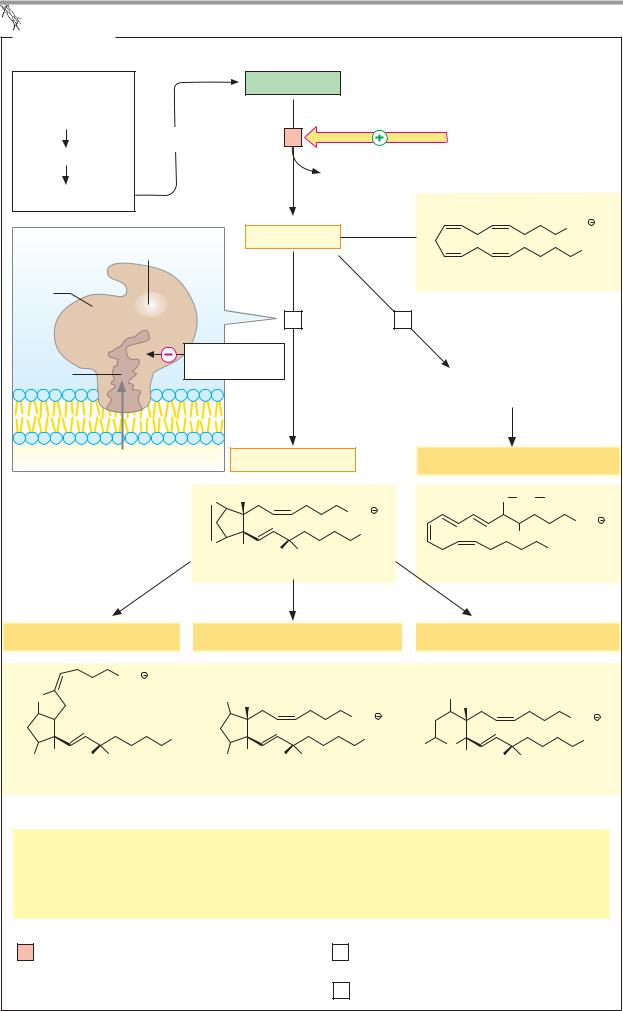
A. Eicosanoids |
|
|
|
|
|
|
|
Essential |
|
|
|
Phospholipid |
|
|
|
fatty acids: |
|
|
|
|
|
|
|
Linoleic acid |
|
|
|
|
|
Hormones and |
|
|
Inclusion in |
|
1 |
|
|
||
Linolenic acid |
|
|
other signals |
|
|||
|
|
|
|
Lysophospholipid |
|
||
|
|
|
|
|
|
||
Arachidonic acid |
|
|
|
|
|
|
|
Prostaglandin |
Peroxidase |
|
Arachidonate |
|
COO |
||
|
|
CH3 |
|||||
synthase |
center |
|
|
|
|
|
|
|
|
|
|
|
|
||
Heme |
|
|
|
|
|
|
|
|
|
|
|
2 |
Prosta- |
3 Lipoxygenase |
|
Cyclooxy- |
|
|
|
|
glandin |
|
|
Ser |
Acetylsalicylic |
synthase |
|
|
|||
genase |
|
|
|||||
|
|
|
|||||
channel |
|
acid |
|
|
|
Hydroxyand |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Hydroperoxy fatty acids |
|
Arachidonate |
|
Prostaglandin H2 |
Leukotrienes |
|
|||
|
|
O |
H |
|
|
S Cys Gly |
|
|
|
|
|
|
COO |
|
COO |
|
|
|
|
|
CH3 |
|
|
|
|
O |
|
|
OH |
|
|
|
|
H |
H |
OH |
CH3 |
|
|
|
|
|
|
||||
|
|
|
Prostaglandin H2 |
Leukotriene D4 |
|||
|
Prostacyclins |
|
|
|
Prostaglandins |
|
|
Thromboxanes |
||
|
|
COO |
|
|
|
|
|
|
|
|
O |
|
|
|
|
HO |
H |
|
|
HO |
|
|
|
|
|
|
|
|
|
H |
|
|
|
|
|
|
|
|
|
COO |
|
COO |
|
|
|
|
|
CH3 |
|
|
|
CH3 |
O |
CH3 |
|
H |
|
|
|
|
H |
|
HO |
|
|
HO |
H OH |
|
|
HO |
H OH |
|
H |
H OH |
||
|
|
|
|
|
|
|||||
|
Prostacyclin I2 |
|
|
|
Prostaglandin F2α |
|
Thromboxane B2 |
|||
|
|
Effects: Stimulation of |
|
|
|
Hormone-controlled lipases |
||||
|
|
|
|
|
|
|
|
|||
|
|
Contraction of smooth muscle |
|
Thrombocyte aggregation |
||||||
|
|
Biosynthesis of steroid hormones |
|
Pain production |
|
|||||
|
|
Gastric juice secretion |
|
|
|
Inflammatory response |
||||
1 |
Phospholipase A |
2 |
3.1.1.4 |
|
|
2 |
Prostaglandin H-synthase [heme] |
|||
|
|
|
|
|
|
|
(dioxygenase + peroxidase) 1.14.99.1 |
|||
3 Arachidonate lipoxygenases 1.13.11. n