




|
|
|
|
|
|
|
|
|
|
Mitochondria |
213 |
||
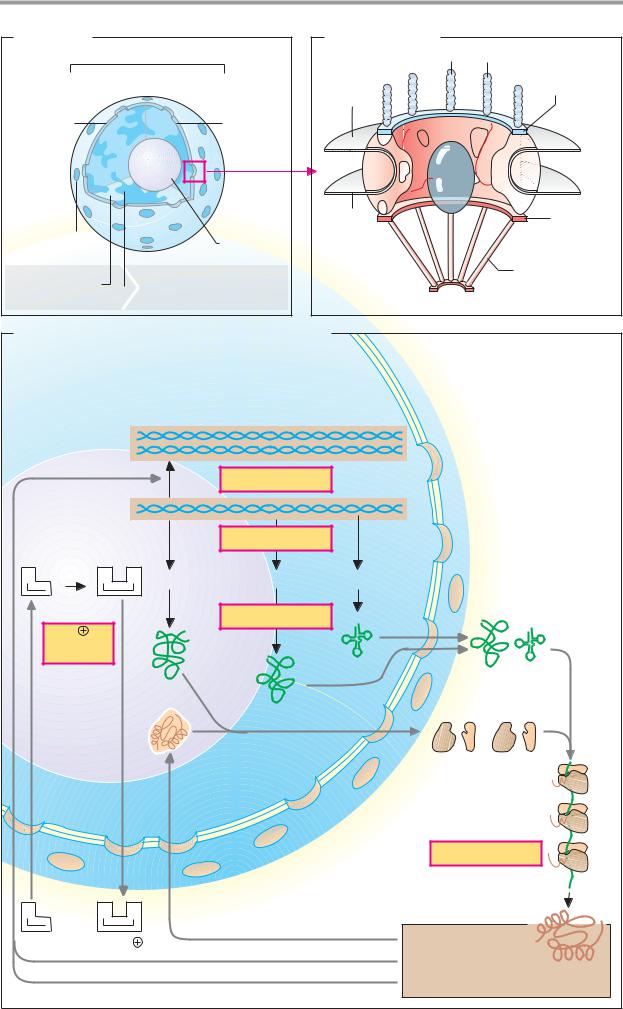
A. Transport systems |
|
|
|
|
|
|
|
|
|
|
|||
Driving force: |
|
ATP 4 |
|
|
|
|
2-Oxoglutarate/citrate |
|
|||||
|
Membrane |
ADP 3 |
|
|
|
|
|
|
|
|
|
|
|
|
potential |
|
|
|
|
|
|
Malate– |
|
|
|||
|
Proton |
|
|
|
|
|
2-oxo- |
|
|
|
|
||
|
gradient |
|
|
|
|
|
glutarate/ |
|
|
|
|
|
|
|
|
|
|
A |
|
|
citrate |
A |
|
|
|
|
|
|
H2PO4 |
|
ATP |
4 |
|
|
|
|
|
2 |
|
||
|
|
|
|
|
|
|
HPO4 |
|
|||||
H2O |
|
|
|
|
|
Malate |
– |
|
|
||||
|
|
|
|
ADP 3 |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
A |
|
|
|
||
|
|
|
|
|
|
|
Citrate export |
|
|
|
|
||
|
OH |
|
H2PO4 |
|
|
|
|
|
|
||||
|
|
|
Malate shuttle |
|
2 |
|
|
|
|||||
H |
+ |
|
A |
|
|
|
|
|
HPO4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
U |
|
|
|
|
|
|
|
OH |
|
ATP synthesis |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
Ca2 + |
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
||
Pyruvate |
|
A |
|
|
|
Ca2+ storage |
Ca2 + |
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
H + |
|||
|
|
|
Pyruvate |
|
Breakdown |
Ca2+ release |
|
|
|
||||
|
|
|
|
Conversion |
|
|
|
|
A |
(Na+ ) |
|||
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
Inner mitochondrial membrane |
|
|
|
|
|
|
||||
B. Malate and glycerophosphate shuttle |
|
|
|
|
|
|
|
||||||
1 Glyceraldehyde |
2a 2b Malate dehydrogenase 1.1.1.37 |
4 Glycerol 3-phosphate dehydrogenase 1.1.1.8 |
|||||||||||
|
3-phosphate |
|
|
|
|
|
|
|
|
|
|
|
|
|
dehydrogenase |
3a 3b Aspartate transaminase 2.6.1.1 |
5 Glyceral 3-phosphate DH (FAD) 1.1.99.5 |
|
|||||||||
|
1.2.1.12 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
NADH + H + |
|
|
|
Glucose 6- |
|
|||
|
|
|
|
|
1,3-Bisphos- |
Glyco- |
phosphate |
|
|||||
Cyto- |
|
|
|
Cytoplasma |
|
|
|
||||||
|
|
|
phoglycerate |
lysis |
|
|
|
||||||
plasm |
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
||
|
Glu |
|
Oxalo- |
|
|
N |
A |
|
|
|
Fructose 1,6- |
||
|
|
|
acetate |
|
|
|
|
1 |
|
|
bisphosphate |
||
|
|
3b |
|
|
2a |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
N |
A |
Glyceral 3- |
|
Glycerone 3- |
|||
|
Asp |
|
2-Oxo- |
|
|
phosphate |
|
phosphate |
|
||||
|
|
Malate |
|
|
|
|
|||||||
|
|
|
glutarate |
|
|
|
|
|
|
4 |
|
||
Outer |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
Glycerophos- |
Glycerol 3- |
|
||
|
|
|
|
|
|
|
|
|
phate shuttle |
phosphate |
|
||
Mitochon- |
Malate |
|
|
|
|
|
|
|
|
|
|
||
drial |
|
|
|
|
|
|
|
|
|
|
|
||
|
shuttle |
|
|
|
|
|
|
|
|
Porin |
|||
membrane |
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
Complex I |
|
|
|
|
|
Inner |
|
|
|
|
|
|
I |
Ubiquinone |
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|||
|
Asp |
|
2-Oxo- |
Malate |
|
|
|
Q |
|
|
|
|
|
|
|
|
|
N |
A |
|
|
|
|
|
|||
|
|
|
glutarate |
|
|
|
|
|
|
|
|
||
|
|
3a |
|
|
2b |
|
|
|
|
|
5 |
|
|
|
|
|
|
|
|
Complex III |
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|||
|
Glu |
|
Oxalo- |
|
|
N |
A |
|
|
|
|
|
|
|
|
acetate |
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
Complex IV |
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|||
Matrix |
|
|
|
NADH + H+ |
2H+, O |
|
|
|
|
|
|||
|
|
|
|
|
Mitochondrion |
|
H2O |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|
|
Biological membranes |
215 |
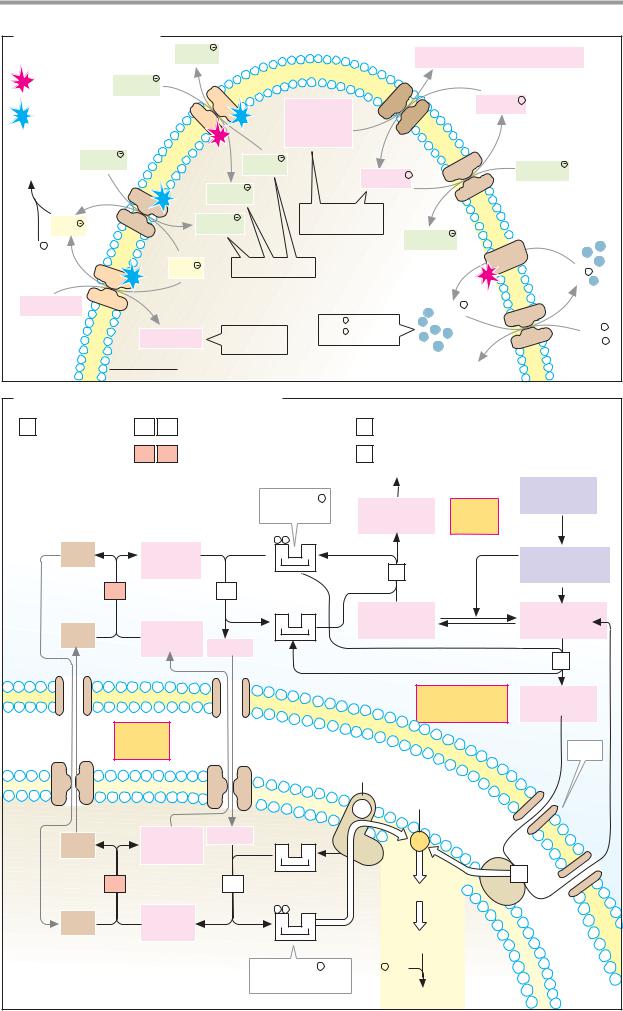
A. Structure of the plasma membrane |
|
|
|
|
|
Extracellular |
|
|
|
side |
|
Glycoprotein |
|
Oligosaccharide |
|
Phospholipid |
|
|
|
|
|
|
|
Lipid |
|
Glycolipid |
|
bilayer |
|
||
|
|
|
|
|
|
|
5 nm |
Peripheral |
|
|
|
membrane |
Integral |
Cytoplasmic |
|
protein |
membrane proteins |
side |
|
B. Membrane lipids
|
|
|
|
|
a |
|
b |
|
c |
|
d |
|
||
a |
Phosphatidyl- |
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
inositol |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Sphingomyelin |
|
|
|
|
|
|
|
|
|
|
|
|
|
b |
Polar |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ganglioside |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
c |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
d |
Phosphatidyl- |
|
|
|
|
|
|
|
|
|
|
|
|
Outer |
|
ethanolamine |
|
|
|
|
|
|
|
|
|
|
|
|
leaflet |
eCerebroside
|
|
Apolar |
|
|
f |
Phosphatidyl- |
|
|
|
|
||||
|
|
|
||
|
choline |
|
|
|
|
|
|
|
Inner |
g |
Cholesterol |
|
|
|
|
|
leaflet |
||
|
|
|
|
hPhosphatidylserine
|
Phospholipids |
Polar |
|
|
|
|
Glycolipids |
|
|
|
|
|
Cholesterol |
e |
f |
g |
h |
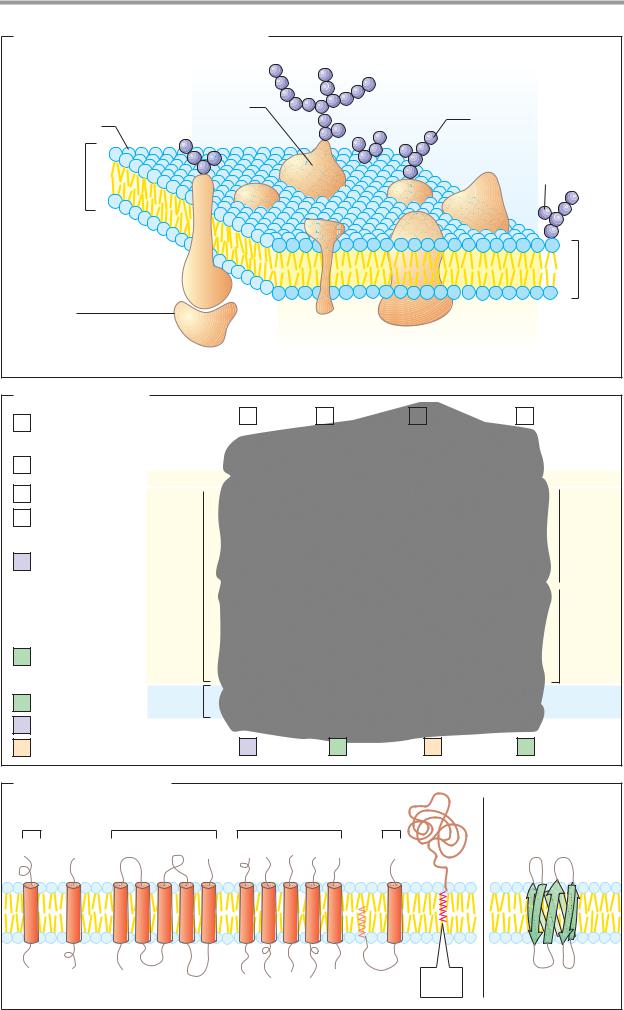
C. Membrane proteins |
|
|
|
||
Type I Type II |
Type III |
Type IV |
Type V |
|
|
N |
C |
|
|
|
|
|
|
|
|
TypeVI |
|
C |
N |
|
|
Lipid |
|
|
|
|
|
|
|
1. α-Helical |
|
|
anchor |
2. β-Barrel |
|


Biological membranes |
217 |
A. Functions of membranes
|
|
|
S |
|
|
|
|
|
A |
B |
|
|
|
|
|
|
|
2 |
3 |
4 |
|
|
|
|
|
|
|
|
|
||
1 |
|
|
|
A+B |
5 |
|
|
|
|
|
|
|
|
||
|
A |
B |
|
|
|
|
|
|
|
C+D |
|
|
6 |
||
|
|
|
|
|
|
|
|
Boundary |
Controlled |
Signal re- |
Enzymatic |
Contact |
|
Anchor |
|
|
metabolite |
ception and |
reactions |
with |
|
for cyto- |
|
|
transport |
transmission |
|
other cells |
|
skeleton |
|
B. Composition of membranes |
|
|
|
|
|||
Membrane components |
|
|
Relative proportion of lipids |
||||
|
|
|
Nerve cell: Plasma membrane |
|
|
|
|
|
|
|
Erythrocyte: Plasma membrane |
|
|
||
|
|
|
|
** |
|
|
|
|
|
|
|
* |
|
|
|
|
|
|
* |
|
|
|
|
|
|
|
|
* |
|
|
|
|
|
|
* |
* |
|
|
|
|
|
|
Liver cell: Plasma membrane |
Cardiolipin |
|
||
|
|
|
|
|
|
||
Inner membrane |
|
Mitochondrion |
|
Both membranes |
|||
|
|
|
|
||||
|
|
|
|
|
Phosphatidyl- |
* |
|
|
Lipids |
|
|
|
choline |
|
|
|
|
|
|
|
Phosphatidyl- |
* |
Glycolipids |
|
Carbohydrates |
|
|
serine |
|||
|
Phospholipids |
|
|||||
|
|
|
|
|
Phosphatidyl- |
|
Cholesterol |
|
Proteins |
|
|
|
ethanolamine * |
||
|
|
|
|
|
|||
|
|
|
|
|
Sphingomyelin * |
Other lipids |
|



|
|
|
Biological membranes |
|
221 |
||
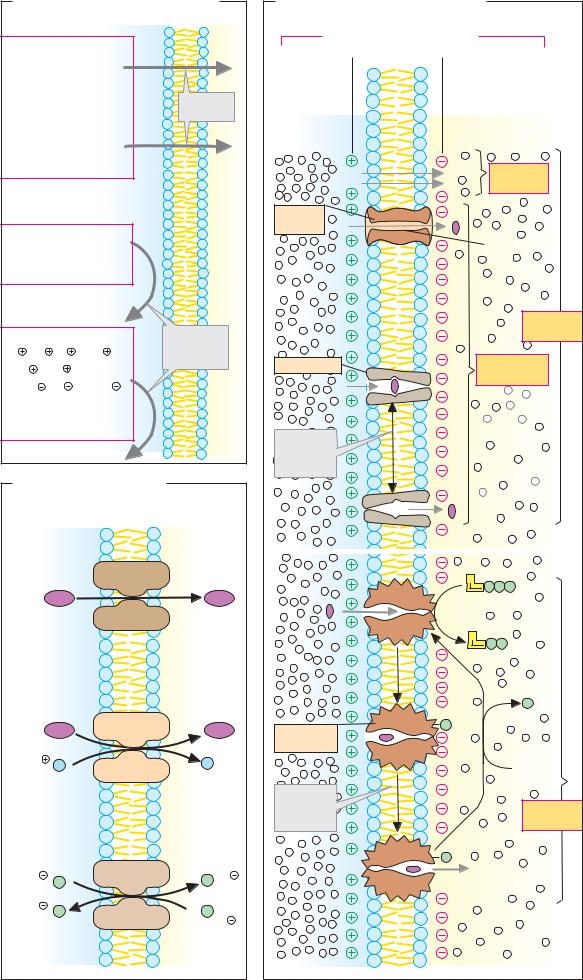
A. Transport mechanisms |
|
|
|
|
Blood |
|
|
|
|
|
Lumen |
Epithelial cells |
|
||
|
|
|
|
|
|
||
N |
|
|
|
A |
|
Na |
|
|
|
|
|
P P |
P |
||
E |
|
|
Na |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
S |
A |
|
|
|
M |
M |
|
|
P |
P |
|
|
+ P |
|
|
+ P |
|
|
||
|
A |
|
|
|
|
|
|
|
|
P P |
|
|
|
|
|
|
A |
P P P |
M |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
M |
|
|
M |
|
|
|
|
U |
|
|
|
|
|
|
|
|
|
|
|
1. Facilitated diffusion |
2. Active transport |
3. Secondary active transport |
|
|
|||
B. Glucose transporter Glut-1 |
|
D. Sarcoplasmic Ca2 pump |
|
|
|||
|
|
|
Binding |
ATP |
|
N |
Domains |
|
|
|
site for |
|
P |
||
|
|
|
ATP |
|
|
||
|
|
|
|
|
|
β |
|
|
|
|
|
|
|
|
|
N |
C |
|
|
|
|
|
|
Glucose |
|
|
|
|
|
|
|
Blood |
|
|
|
|
“Stalk” |
|
|
|
|
|
Cyto- |
|
|
|
|
Plasma |
|
|
plasm |
|
|
|
|
|
|
|
|
|
|
|
|
membrane |
|
|
|
|
|
|
|
|
|
|
Binding |
|
Trans- |
|
|
|
|
|
sites |
|
membrane |
||
|
|
|
for Ca2 |
|
domain |
||
Cytoplasm |
|
|
|
|
|
|
|
|
Glucose |
|
SR |
|
|
|
|
|
|
1. Structure |
|
|
|
|
|
|
|
|
|
|
|
|
|
C. Aquaporin-1 |
|
|
|
A |
|
|
|
|
|
|
P P P |
|
|
|
|
|
|
|
A |
|
Ca2 |
|
|
|
|
|
P P P |
|
|
|
|
|
|
|
|
|
A |
|
|
|
|
|
|
|
P |
P |
|
|
|
|
|
a) |
P |
|
|
|
|
|
|
|
|
||
N |
C |
|
|
|
|
|
|
Tubular |
H2O |
|
|
|
|
|
|
lumen |
|
d) |
P |
b) |
|
|
|
|
|
|
|
|
|||
Plasma |
|
|
|
c) |
|
|
|
membrane |
|
|
|
|
|
|
|
|
|
|
P |
|
A |
|
|
|
|
|
|
P P |
|
|
|
|
|
|
|
|
|
|
|
Tubule cell |
|
|
2. Catalytic cycle |
|
|
|
|

|
|
|
|
Biological membranes |
223 |
|
A. Voltage-gated Na+ channel |
|
|
|
|||
I |
II |
|
III |
IV |
|
|
|
|
|
|
2 |
3 |
|
|
|
|
|
1 |
4 |
|
|
1 2 3 4 5 |
6 |
|
6 |
5 |
|
N |
|
|
|
C |
Membrane |
|
|
|
|
|
a |
||
|
|
|
|
Narrow pore |
polarized, |
|
|
|
|
|
|
helix 4 |
|
|
Pore |
|
α-Subunit |
Na |
in position 1 |
|
|
|
|
|
|
||
|
|
|
|
|
4 |
|
|
|
|
|
|
3 |
|
Voltage- |
|
|
|
2 |
|
|
|
|
|
Outside |
5 |
|
|
sensitive |
|
|
|
1 |
|
|
|
|
|
6 |
|
|
|
helix |
|
|
|
|
|
|
(helix 4) |
|
|
|
|
|
|
Inside |
b |
Membrane |
|
Wide pore |
depolarized, |
|
helix 4 |
|
1. Structure |
2. Mechanism |
in position 2 |
Na |
B. Nicotinic acetylcholine receptor |
|
|
|
|
|||
N α C |
δ |
β |
α |
γ |
δ |
β |
αH |
|
|
|
|
|
α L |
|
|
|
|
|
|
|
|
|
Acetyl- |
|
|
|
|
|
|
|
choline |
|
|
|
Subunits |
|
|
|
Na |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Helix 2 |
1. Structure |
|
|
|
2. Pore |
|
3. Mechanism |
|
C. K + |
channel in Streptomyces lividans |
|
|
|
|||
A |
B |
|
C |
D |
|
|
Outside |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Plasma- |
|
|
|
|
|
|
|
membrane |
|
|
|
Subunits |
|
|
|
|
|
|
Outside |
|
|
|
|
|
|
|
Plasma |
|
|
Inside |
|
|
|
|
|
|
|
|
|
|
|
|
membrane |
|
|
|
|
|
|
|
Inside |
|
|
K |
|
|
|
|
|
|
|
|
|
|
|
Homotetramer |
|
|
|
From above |
||

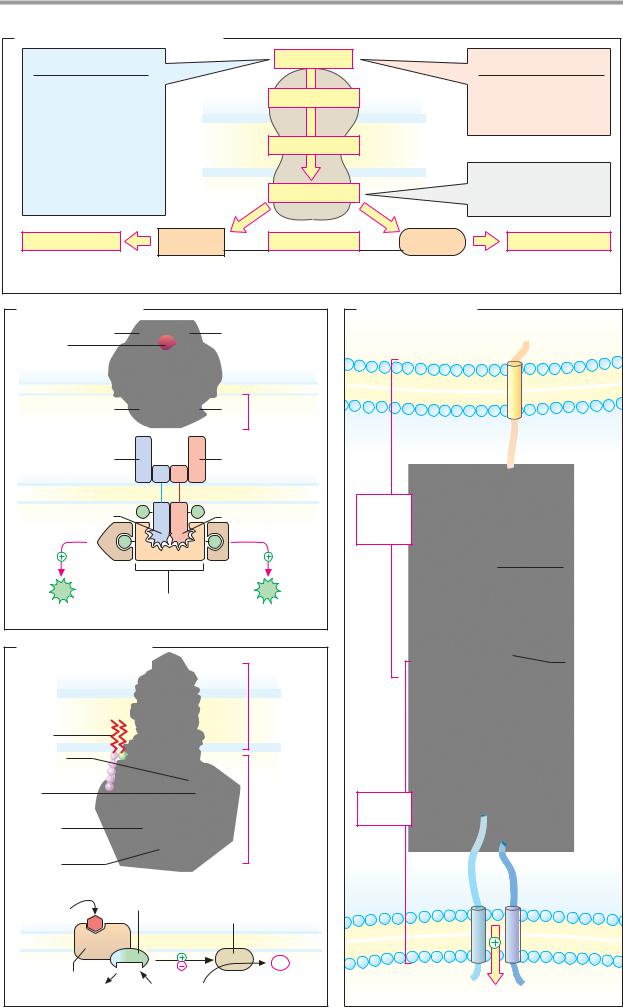



|
|
|
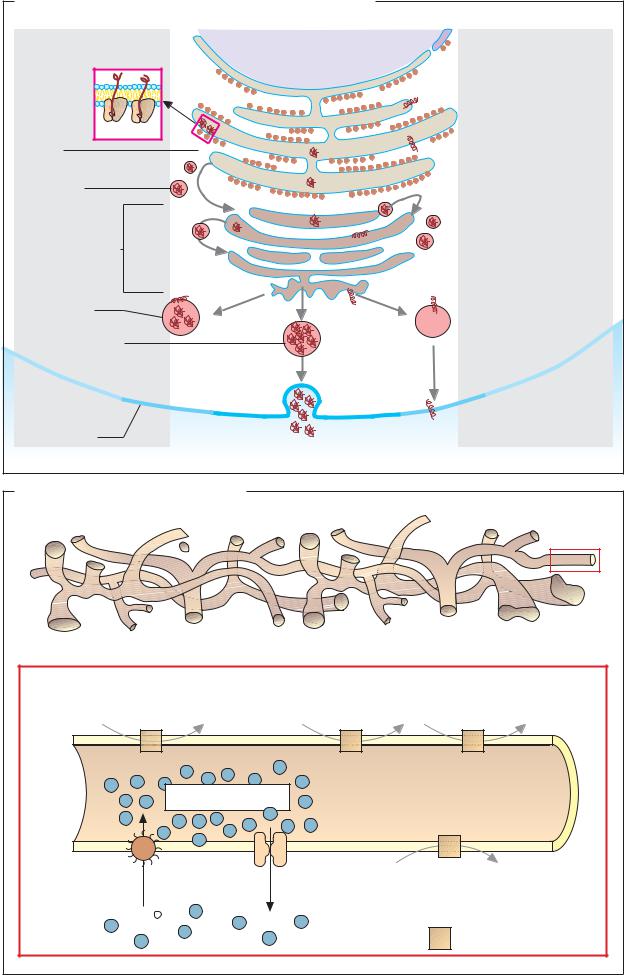
Endoplasmic reticulum and Golgi apparatus |
229 |
||||||
A. Protein sorting |
|
|
|
|
|
|
|
|
|
|
Cytoplasmic pathway |
Ribosomes |
Secretory pathway |
|
|
|
|||||
|
|
|
|
4 |
|
2 |
|
|||
|
|
|
* |
+ H |
N + + |
|
|
-KDEL |
||
|
|
Protein |
|
|
|
|
|
|||
* |
|
|
3 |
1 |
|
Rough ER |
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
Retention |
Cytoplasm |
|
|
|
|
|
|
|
|
|
|
-SKL |
5 |
|
|
|
|
* |
|
Retention |
|
|
H2N |
|
|
|
|
Golgi complex |
|
|
|
|
7 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6 |
Receptor |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
R |
|
R |
|
|
|
|
8 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Peroxisomes |
|
Mitochondria |
|
|
3 |
* |
Secretory |
|
||
|
|
R |
|
|
|
|
vesicle |
|
||
|
|
|
|
|
|
|
|
|||
|
Nucleus |
|
|
Lysosomes |
|
9 |
Ca |
2 + |
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cell membrane |
|
|
|
|
B. Translocation signals |
|
|
|
*Standard pathway (without signal) |
||||
|
|
|
C. Exocytosis |
|
|
|
||
|
Signal peptide |
|
+ + |
α-Helix |
Transmitter |
Vesicle |
Rab · GTP |
|
1 |
+ H2N |
|
||||||
(secretory pathway) |
|
|
|
|
|
|
|
|
|
|
+ OOC |
|
|
Synapto- |
|
|
GTP |
|
|
|
|
|
|
|
||
|
|
|
|
|
brevin |
|
|
|
2 |
Signal sequence |
|
|
|
(v-SNARE) |
|
|
Synapto- |
(ER proteins) |
|
...LEDK.... |
Syntaxin |
|
|
tagmin |
||
|
|
|
|
|
|
|||
|
|
|
Mannose 6- |
(t-SNARE) |
|
|
|
|
|
Signal group |
|
|
|
|
|
||
3 |
|
phosphate |
|
|
|
Ca2 |
||
(lysosomal proteins) |
|
|
P |
Synaptic cleft |
|
|
||
|
|
|
|
|
1. |
|
|
Channel |
|
|
|
|
|
|
|
|
|
|
|
|
|
Apolar |
|
|
|
GTP |
4 |
Stop-transfer sequence |
|
sequence |
|
|
|
||
|
|
|
|
|
||||
(membrane proteins) |
|
|
|
|
|
|
||
|
|
|
+ + + |
|
Synaptotagmin |
|
|
|
|
Signal peptide |
|
|
releases |
Ca |
2 |
Action |
|
|
|
|
|
v-SNAREs |
potential |
|||
5 |
(mitochondrial |
|
|
|
|
|||
|
|
|
|
|
|
|
||
|
proteins) |
|
|
|
|
|
|
|
|
|
|
|
...KKKRK... |
2. |
|
|
Ca2 |
|
Signal sequence |
|
|
|
|
|
||
6 |
|
|
|
|
|
|
Rab · GDP |
|
(nuclear proteins) |
|
|
|
|
|
|
||
|
Signal sequence |
|
|
|
|
|
|
GDP |
7 |
|
|
|
Membrane |
|
|
P |
|
(peroxisomes) |
|
...FKS... |
|
|
||||
|
|
|
fusion |
|
|
SNAP |
||
|
|
|
|
|
|
|
|
+NSF |
8 |
Signal region |
|
|
|
|
|
|
|
(secretory vesicle) |
|
|
|
3. |
SNARE complex |
|||
|
|
|
|
|
||||


Endoplasmic reticulum and Golgi apparatus |
231 |
A. Protein synthesis in the rough endoplasmic reticulum |
|
|
|
1 |
2 |
3 |
4 |
mRNA |
SRP |
|
|
|
|
|
|
Signal |
|
|
|
peptide |
|
|
|
Free |
|
|
|
ribosome |
|
|
|
ER membrane |
|
|
|
ER-lumen |
SRP receptor |
Translocon |
|
5 |
6 |
7 |
8 |
GDP+Pi |
|
|
|
GTP |
|
|
|
Signal peptidase |
|
|
Post-transl. |
|
|
modification |
|
B. Protein glycosylation
Protein
1 glycosyltransferase
2.4.1.119
Dolichol (Dol) |
Ribosome |
2 O-glycosidases 3.2.1.n |
Translocon |
3 Glycosyltransferases
2.4.1.n
P |
Glucose |
|
1 |
||
|
||
P |
N-acetylglucosamine |
|
Asn |
||
|
Mannose |
|
Asn |
Galactose |
|
|
N-acetylneuraminic |
|
|
acid |
UDP |
H2O |
|
|
||
GDP |
Asn |
|
Dol |
2 |
|
Precursors |
||
Dol |
||
|
||
Core structure |
Asn |
|
P |
3 |
|
P |
Mannose-rich |
|
type |
||
|
Dolichol diphosphate (9–23 isoprene units) |
Complex |
|
type |
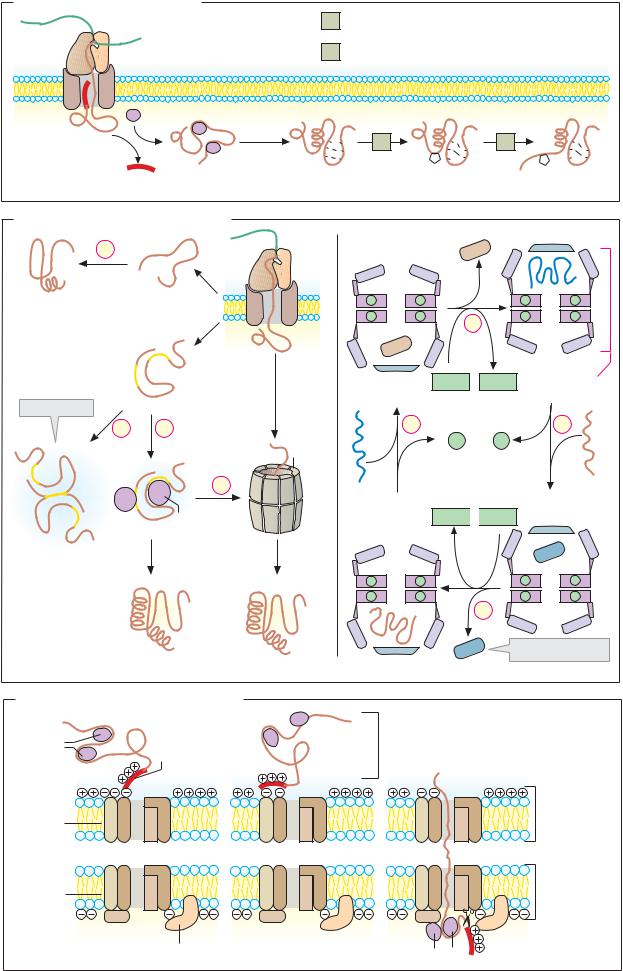

 7 ATP
7 ATP