


Basics 197
A. Comparison of prokaryotes and eukaryotes
|
Prokaryotes |
|
Eukaryotes |
|
|
Organisms |
|
|
|
1-10 m |
Eubacteria |
|
Fungi |
|
Archaebacteria |
|
Plants |
|
|
|
|
|
Animals |
|
|
|
Form |
|
|
|
Single-celled |
Single or multi-cellular |
|
|
|
Organelles, cytoskeleton, cell division apparatus |
10-100 m |
||
|
Missing |
|
Present, complicated, specialized |
|
Small, circular, no |
DNA |
|
|
|
introns, plasmids |
|
Large, in nucleus, many introns |
||
|
RNA: Synthesis and maturation |
|
||
Simple, in cytoplasm |
|
Complicated, in nucleus |
|
|
|
Protein: Synthesis and maturation |
|
||
|
Simple, coupled with |
|
Complicated, in the cytoplasm |
|
|
RNA synthesis |
|
and the rough endoplasmic reticulum |
|
|
Metabolism |
|
|
|
|
Anaerobic or aerobic |
|
|
|
|
very flexible |
|
Mostly aerobic, compartmented |
|
Endocytosis and Exocytosis
|
|
no |
yes |
|
|
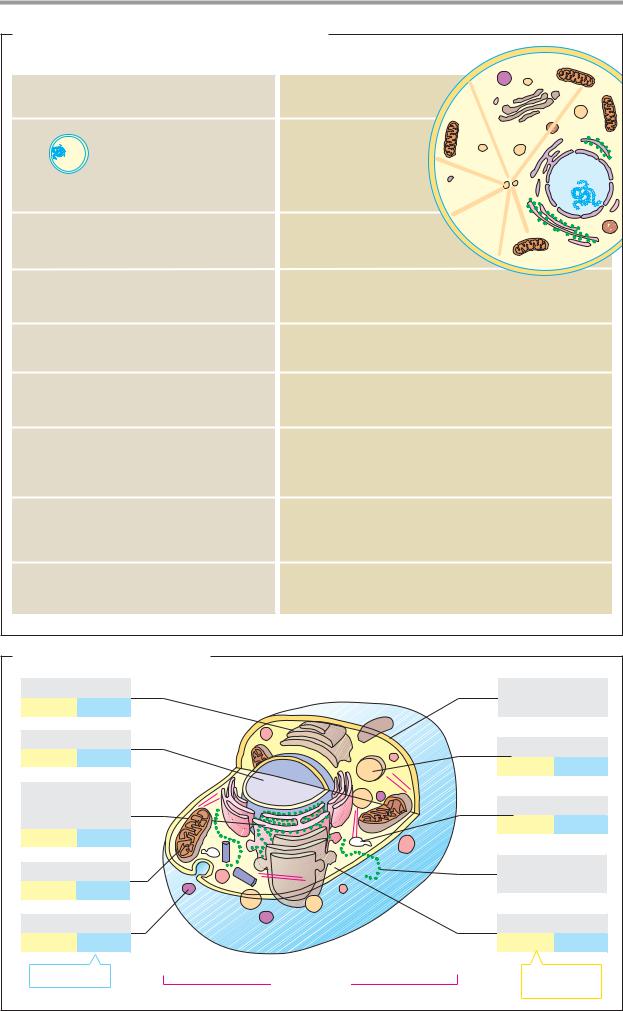
B. Structure of an animal cell |
|
|
|
|
|
Golgi complex |
|
|
Plasma |
|
|
6% |
? |
|
|
membrane |
|
Nucleus |
|
|
|
Lysosome |
|
6% |
1 |
|
|
||
|
|
1% |
300 |
||
|
|
|
|
||
Rough |
|
|
|
|
|
endoplasmic |
|
|
Endosome |
||
reticulum |
|
|
1% |
200 |
|
9% |
1 |
|
|
||
|
|
|
|
||
Mitochondrion |
|
|
Free |
|
|
|
|
ribosomes |
|||
22% |
~2000 |
|
|
||
|
|
|
|
||
Peroxisome |
|
|
Cytoplasm |
||
1% |
400 |
|
|
54% |
1 |
Number per cell |
10-30 |
m |
Proportion of |
||
|
|
cell volume |
|||
|
|
|
|
||

|
|
|
Basics |
199 |
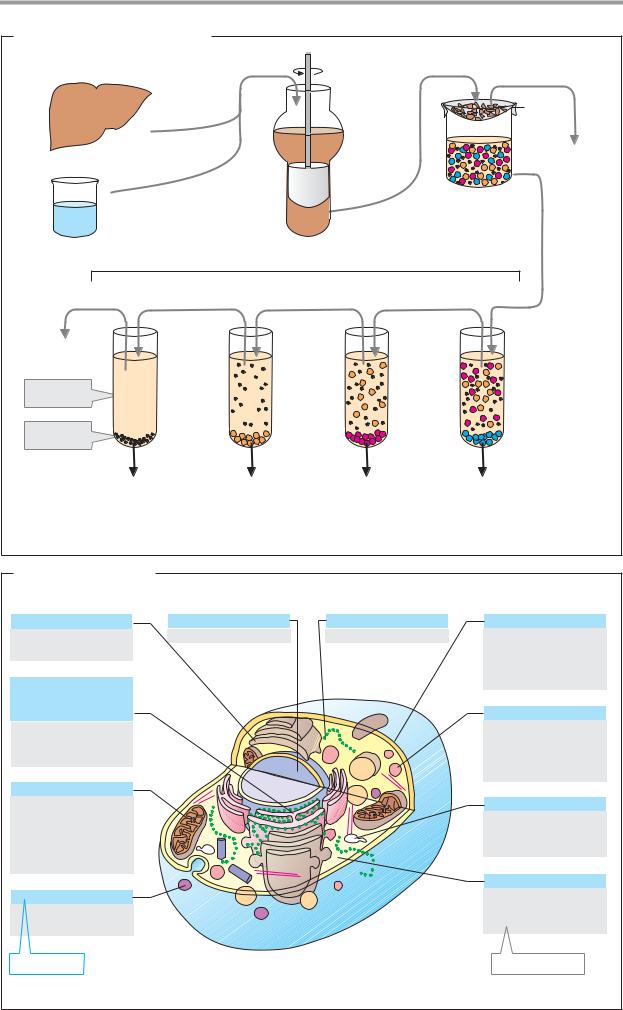
A. Isolation of cell organelles |
|
|
|
|
|
|
Homo- |
Filter |
|
|
|
genize |
|
|
Slice |
|
|
Gauze |
|
|
|
|
|
|
Tissue |
|
|
Whole |
|
|
|
cells, |
|
|
|
|
|
|
|
|
|
|
connective |
|
|
Potter |
|
tissue |
|
|
|
|
|
|
Buffer |
homo- |
|
|
|
genizer |
|
|
|
|
|
|
|
|
|
|
Centrifuge |
|
|
|
g = 300 000 |
g = 100 000 |
g = 15 000 |
g = 600 |
|
120' |
60' |
15' |
10' |
|
Cytosol |
|
|
|
|
Super- |
|
|
|
|
natant |
|
|
|
|
|
|
|
Centrifuge |
|
Pellet |
|
|
tube |
|
|
|
|
|
|
Ribosomes |
Plasma membrane |
Mitochondria |
Nucleus |
|
Viruses |
ER fragments |
Lysosomes |
Cytoskeleton |
|
Macromolecules |
Small vesicles |
Peroxisomes |
|
|
|
Microsomal |
(Plants: |
|
|
|
fraction |
chloroplasts) |
|
|
B. Marker molecules |
|
|
|
Golgi complex |
Nucleus |
Ribosomes |
Plasma membrane |
α-Mannosidase II |
DNA |
rRNA |
Na+/K+ ATPase |
3.2.1.24 |
|
|
3.6.1.37 |
|
|
|
Phosphodiesterase I |
Endoplasmic |
|
|
3.1.4.1 |
|
|
|
|
reticulum |
|
|
Lysosome |
|
|
|
|
Glucose 6-phospha- |
|
|
β-N-Acetylhexos- |
tase 3.1.3.9 |
|
|
aminidase 3.2.1.52 |
RNA |
|
|
β-Galactosidase |
|
|
|
3.2.1.23 |
Mitochondrion |
|
|
|
Succinate dehydro- |
|
|
Endosome |
genase 1.3.5.1 |
|
|
Uptake of |
Cytochrome-c |
|
|
peroxidase |
oxidase |
|
|
1.11.1.7 |
1.9.3.1 |
|
|
Cytosol |
|
|
|
|
Peroxisome |
|
|
L-Lactate |
Catalase |
|
|
dehydrogenase |
1.11.1.6 |
|
|
1.1.1.27 |
Cell fraction |
|
|
Marker enzyme |


|
|
|
Basics |
203 |
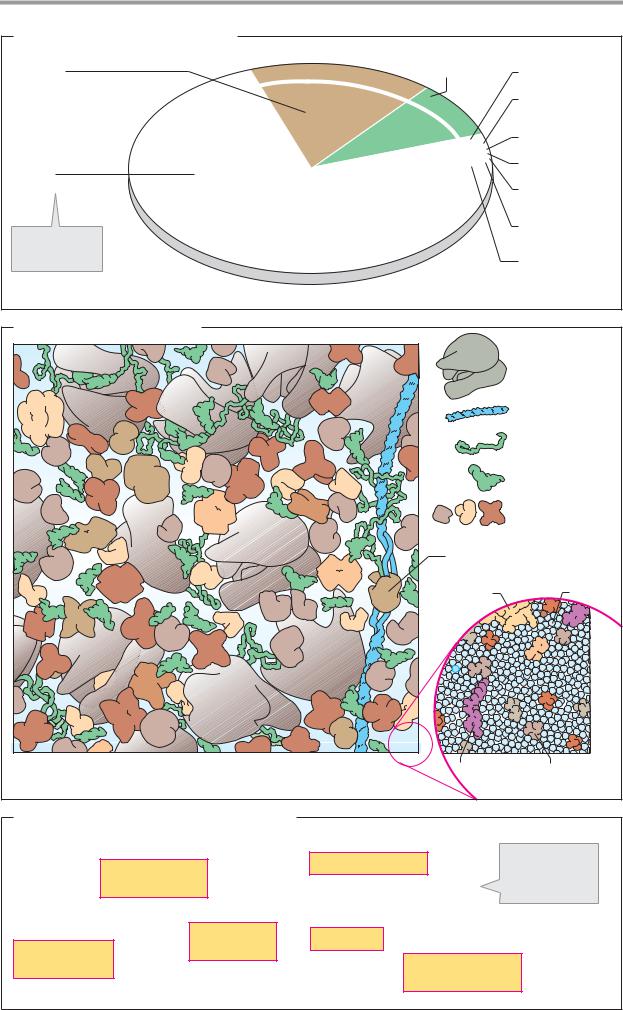
A. Components of a bacterial cell |
|
|
|
|
Proteins |
M |
RNA (8%) |
DNA (1%) |
|
|
|
|
|
|
(17%, 3 000) |
ac |
romol |
|
|
|
|
e |
|
|
|
|
c |
|
|
|
|
ul |
|
|
|
|
e |
Sugars (1%, 250) |
|
|
|
s |
||
|
|
|
Inorganic |
|
|
|
|
ions (1%, 20) |
|
Water |
|
|
Lipids (1%, 50) |
|
|
|
|
|
|
(70%, 1) |
|
|
Nucleotides |
|
|
|
|
(0.4%, 100) |
|
|
|
|
Amino acids |
|
Number of |
|
|
(0.4%, 100) |
|
different mole- |
|
|
Other small |
|
cule types |
|
|
|
|
|
|
|
organic |
|
|
|
|
molecules |
|
|
|
|
(0.2%, 300) |
|
B. View into a bacterial cell
Ribosome
DNA
mRNA
tRNA
Proteins
RNA-Polymerase
Protein |
Water |
|
|
|
Carbohydrate |
Amino acid |
C. Biochemical functions of the cytoplasm |
|
|
|
|
|
|
Gluconeogenesis |
Schematic |
|
|
Pentose phos- |
net of the |
||
|
|
|||
|
phate cycle |
|
reactions |
|
|
|
in the cytoplasm |
||
|
|
|
||
|
Fatty acid |
Glycolysis |
|
|
Protein |
biosynthesis |
And many other |
|
|
biosynthesis |
|
|
||
|
|
reactions |
|
|