




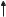

 Protein synthesis
Protein synthesis 
 Blut-Glucose
Blut-Glucose 

|
|
|
|
|
|
Lipophilic hormones |
377 |
|||
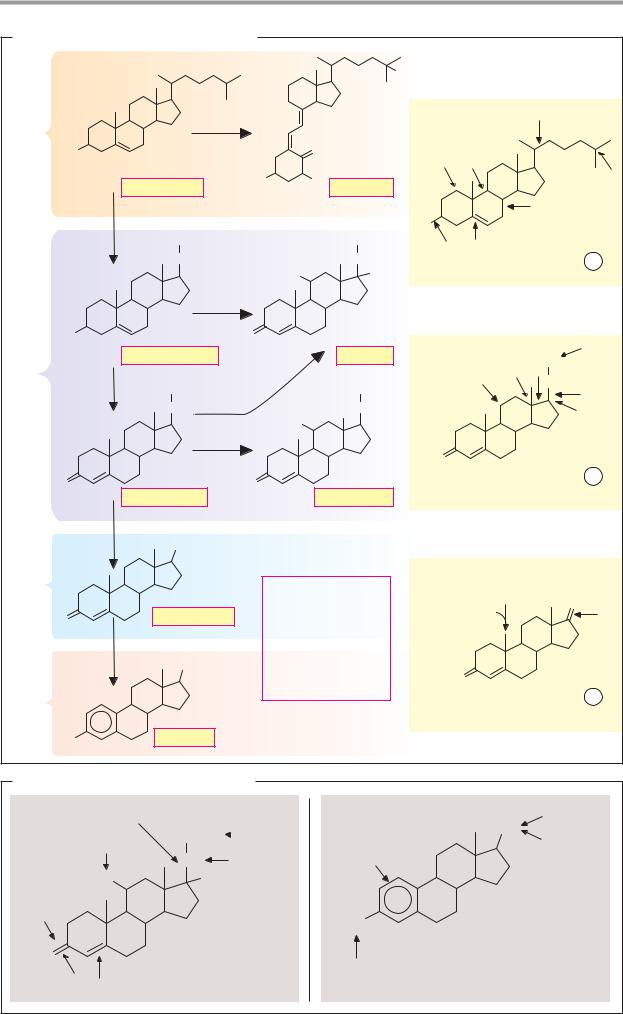
A. Biosynthesis of steroid hormones |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
OH |
|
|
|
|
|
|
DSHH |
|
|
|
|
|
H |
|
|
C27 |
|
|
|
|
|
|
|
|
|
|
HO |
|
|
CH2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
Cholesterol |
HO |
OH Calcitriol |
|
C |
D |
|
H |
||
|
|
|
|
|||||||
|
H |
|
|
|
|
A |
B |
D |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H |
CH3 |
|
|
CH2OH |
HO |
|
|
|
|
|
H/S |
|
|
D |
I |
|
|
|
||
|
|
CO |
|
|
CO |
|
|
1 |
||
|
|
|
|
Cholesterol |
|
|
||||
|
|
|
HO |
|
OH |
|
|
|||
|
|
|
|
|
|
|
|
|
||
|
|
HYDHH |
|
|
|
|
|
|
|
|
|
HO |
|
O |
|
|
|
|
|
|
|
C21 |
Pregnenolone |
|
Cortisol |
|
H |
D |
CH3 |
|
||
|
|
|
|
|
|
|
||||
Y |
|
|
|
|
|
|
|
|||
CH3 |
|
|
CH2OH |
H |
|
CO |
H |
|||
|
D |
|
|
|
|
|
|
|||
|
|
CO HHH |
HO |
OHC |
CO |
|
C |
D |
S |
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
HHHD |
|
|
|
A |
B |
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
O |
|
O |
|
|
Progesterone |
|
|
2 |
|
|
|
|
|
|
|
|
||||
|
Progesterone |
|
Aldosterone |
|
|
|
||||
|
|
|
|
|
|
|
||||
H
S
YOH
C19 |
|
H: Hydroxylation |
|
A |
|
|
D: Dehydrogenation |
H |
O |
|
Progesterone |
Y |
||
O |
|
|||
I : Isomerization |
|
|
||
|
H |
|
C D |
|
|
Y: Hydrogenation |
|
||
|
A |
A |
B |
|
|
OH |
S: Cleavage |
||
|
|
A: Aromatization |
O |
|
|
|
|
|
|
|
|
|
3 |
C18 |
|
|
|
Androstenedione |
|
|
|
|
|
HO |
Estradiol |
|
|
|
B. Inactivation of steroid hormones |
|
|
||
Oxidative cleavage |
Conjugate |
|
Oxidation |
|
|
CH2OH |
formation |
|
OH |
Oxidation |
|
|
Conjugate |
|
|
CO |
Reduction |
Hydroxylation |
formation |
|
|
|||
|
|
|
||
HO |
OH |
|
|
|
Conjugate |
|
|
|
|
formation |
|
|
HO |
|
|
|
|
|
|
O |
|
|
Conjugate formation |
|
|
|
|
||
Reduction |
Cortisol |
|
Estradiol |
|

|
|
|
|
Lipophilic hormones |
379 |
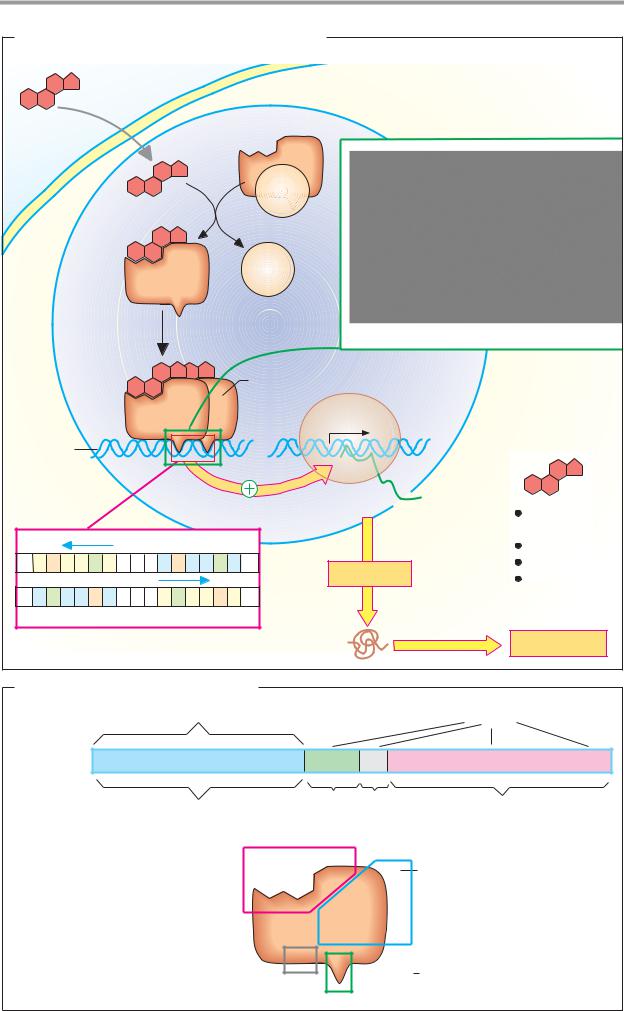
A. Mechanism of action of lipophilic hormones |
|
|
|||
Hormone |
|
|
Target cell |
|
|
|
|
Hormone |
|
|
|
|
|
receptor |
|
|
|
|
|
hsp |
|
|
|
|
|
90 |
|
|
|
|
|
hsp |
|
|
|
|
|
90 |
|
|
|
|
|
Heat-shock |
|
|
|
Nucleus |
2x |
protein |
Glucocorticoid receptor/DNA complex |
||
|
|||||
|
|
Hormone-receptor |
DNA-binding |
|
|
|
|
domain (dimer) |
|
||
|
|
dimer |
RNA |
bound to DNA |
|
|
|
|
|
|
|
|
|
|
polymerase |
Gene |
|
DNA |
|
|
|
|
|
|
|
|
mRNA |
|
|
|
|
|
|
Steroid |
|
|
|
|
|
hormone |
|
|
|
DNA |
|
T3, T4 |
|
A G A A C A n n n |
T G T |
T C T |
Translation |
Calcitriol |
|
|
|
|
Retinoic acid |
||
T C T T G T n n n A C A A G A
Hormone response element (HRE)
|
|
Protein |
|
Cell response |
|
B. Receptors of lipophilic hormones |
|
|
|||
|
Variable length |
|
|
Domains |
|
Receptor |
A/B |
C |
D |
E |
|
gene |
|||||
|
|
|
|
||
|
Regulatory |
DNA- |
Nuclear- |
Hormone- |
|
|
domain |
binding |
targeting |
binding |
|
|
|
domain |
sequence |
domain |
|
|
|
Binds ligand |
|
Interaction with other |
|
|
|
|
|
||
|
|
|
|
nuclear components |
|
Receptor protein |
Domain E |
|
|
|
|
~250 aa |
|
|
|
||
with a total of |
|
|
Domain A/B |
||
400 – 1000 aa |
|
|
|
100 – 600 aa |
|
|
|
Domain D |
Domain C Binds to DNA |
||
|
|
~70 aa |
|
||