

7.2. Модели улова на единицу пополнения промыслового стада в дискретной форме
7.2.1. В дискретной форме можно записать практически все известные модификации уравнений Баранова-Бивертона-Холта. Кроме того, в последнее время появились новые дискретные уравнения для уравновешенного состояния запасов, которые не ограничивают закономерности роста, допускают любой закон растянутости пополнения, изменения с возрастом естественной и промысловой смертности.
Ниже записано одно из наиболее подробных уравнений такого вида:
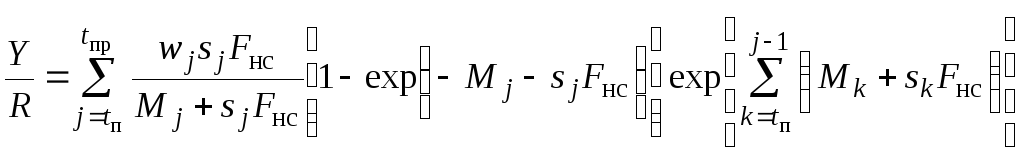 , (7.3)
, (7.3)
где
![]() - возраст вступления рыбы в промысел;
- возраст вступления рыбы в промысел;![]() - предельный возраст рыбы в уловах;
- предельный возраст рыбы в уловах;![]() - коэффициент естественной смертности
рыбы в возрасте
- коэффициент естественной смертности
рыбы в возрасте![]() ;
;![]() - ордината кривой селективности для
рыбы в возрасте
- ордината кривой селективности для
рыбы в возрасте![]() ;
;![]() - коэффициент промысловой смертности
при условно неселективном лове.
- коэффициент промысловой смертности
при условно неселективном лове.
Уравнение (7.3) во многом соответствует уравнению (7.1), записанному в непрерывной форме.
7.2.2.
Ранее в п. 7.1 и 7.2 рассмотрены зависимости
улова и улова на единицу пополнения.
Кроме них, известны уравнения равновесных
продукционных кривых, характеризующих
величину нерестовой части запаса также
на единицу пополнения промыслового
стада
![]() .
.
Первая зависимость характеризует продукционную способность запаса (способность обеспечить некоторую массу нерестового запаса с единицы пополнения), а вторая - относительную величину нерестового запаса, необходимого для поддержания запаса при заданном уровне интенсивности промысла. Ниже приведена одна из дискретных расчетных формул для оценки этой величины с учетом селективного представления промысловой смертности:
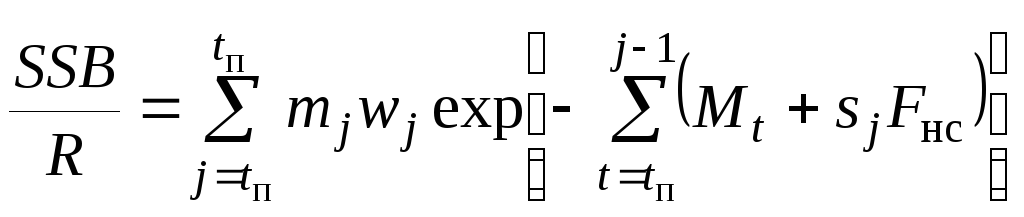 . (7.4)
. (7.4)
Графики
функции
![]() строят в зависимости от коэффициента
мгновенной промысловой смертности так
же, как функции
строят в зависимости от коэффициента
мгновенной промысловой смертности так
же, как функции![]() .
.
Для
решения некоторых задач иногда строят
зависимость
![]() ,
которая показывает, какое количество
рыб пополнения приходится на единицу
нерестовой биомассы. Исследование этой
функции позволяет оценить, до какого
уровня следует увеличивать
,
которая показывает, какое количество
рыб пополнения приходится на единицу
нерестовой биомассы. Исследование этой
функции позволяет оценить, до какого
уровня следует увеличивать![]() для стабилизации запаса при низком
выживании поколения при различной
интенсивности промысла.
для стабилизации запаса при низком
выживании поколения при различной
интенсивности промысла.
7.3. Модели для оценки использования биомассы поколения промысловых рыб
7.3.1.
Важно знать, насколько эффективно
используется биомасса промысловых рыб
при том или ином уровне интенсивности
и селективности рыболовства. При
уравновешенном состоянии запасов и
промысла для этой цели можно использовать
коэффициент
![]() использования биомассы поколения
промысловых рыб. Коэффициент равен
отношению улова рыб одного поколения
к максимально возможной массе этого
поколения в некотором возрасте без
учета промысловой смертности.
использования биомассы поколения
промысловых рыб. Коэффициент равен
отношению улова рыб одного поколения
к максимально возможной массе этого
поколения в некотором возрасте без
учета промысловой смертности.
Математическая
модель для коэффициента
![]() имеет вид:
имеет вид:
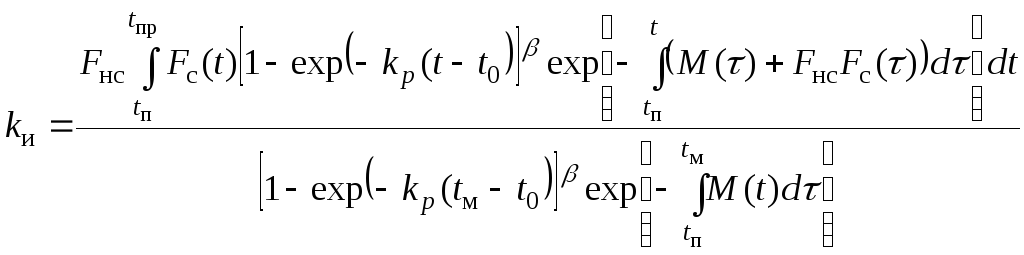 , (7.5)
, (7.5)
где
![]() - возраст, в котором биомасса поколения
достигает максимума.
- возраст, в котором биомасса поколения
достигает максимума.
Остальные условные обозначения приняты такими же, как в выражении (7.1).
В
возрасте
![]() коэффициент естественной смертности
коэффициент естественной смертности![]() часто принимают постоянной величиной.
При постоянном
часто принимают постоянной величиной.
При постоянном![]() и росте рыбы в соответствии с уравнением
Берталанфи возраст
и росте рыбы в соответствии с уравнением
Берталанфи возраст![]() определяют формуле Катти:
определяют формуле Катти:
![]() . (7.6)
. (7.6)
7.3.2.На
примере промысла тихоокеанского хека
определим степень влияния коэффициента
промысловой смертности
![]() при условно неселективном лове и
внутреннего размера ячеи
при условно неселективном лове и
внутреннего размера ячеи![]() сетного мешка или слива орудия лова на
сетного мешка или слива орудия лова на![]() .
.
Сначала
определяют параметры и функции в
выражении (7.5). Особое внимание обращают
на функцию кривой селективности
![]() для различного размера ячеи (см. гл. 3).
для различного размера ячеи (см. гл. 3).
Будем
считать предельный возраст рыбы равным
15 годам, коэффициент естественной
смертности
![]() .
Тогда возраст
.
Тогда возраст![]() согласно (7.6) равен 7,4 года, а максимум
биомассы, отнесенный к единице пополнения,
равен
согласно (7.6) равен 7,4 года, а максимум
биомассы, отнесенный к единице пополнения,
равен![]() г.
г.
В
соответствии с (7.5) коэффициент
![]() является функцией
является функцией![]() и
и![]() .
Графическая иллюстрация этой
зависимости представлена на рис. 7.5 виде
изоплетной диаграммы. На ней по оси
абсцисс отложены значения коэффициента
промысловой смертности
.
Графическая иллюстрация этой
зависимости представлена на рис. 7.5 виде
изоплетной диаграммы. На ней по оси
абсцисс отложены значения коэффициента
промысловой смертности![]() при условно неселективном лове, а по
оси ординат - внутренний размер ячеи
при условно неселективном лове, а по
оси ординат - внутренний размер ячеи![]() .
Наибольших значений коэффициент
.
Наибольших значений коэффициент![]() достигает при
достигает при![]() и
и![]() мм
(на рисунке – штриховая линия).
мм
(на рисунке – штриховая линия).
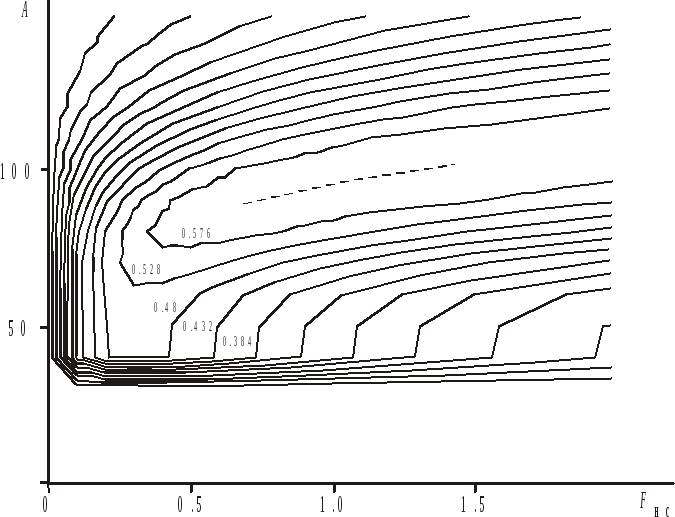
Рис. 7.5. Изоплетная
диаграмма коэффициента использования
биомассы поколения тихоокеанского хека
в зависимости от коэффициента промысловой
смертности
![]() при условно неселективном лове и размера
ячеи тралового мешка
при условно неселективном лове и размера
ячеи тралового мешка
![]() .
.
Оптимальные
значения
![]() и
и![]() определяют, анализируя влияние каждого
из этих факторов на
определяют, анализируя влияние каждого
из этих факторов на![]() .
На рис. 7.6 показаны кривые зависимости
коэффициента
.
На рис. 7.6 показаны кривые зависимости
коэффициента![]() от внутреннего размера ячеи при
фиксированных значениях коэффициента
промысловой смертности
от внутреннего размера ячеи при
фиксированных значениях коэффициента
промысловой смертности![]() .
При увеличении размера ячеи коэффициент
.
При увеличении размера ячеи коэффициент![]() сначала растет и достигает максимума,
равного 0.62–0.63, при размере ячеи 80–100мм
в зависимости от интенсивности промысла.
При этом значения коэффициента
сначала растет и достигает максимума,
равного 0.62–0.63, при размере ячеи 80–100мм
в зависимости от интенсивности промысла.
При этом значения коэффициента![]() расположены в интервале 0.7-1.4.
расположены в интервале 0.7-1.4.
На
рис. 7.7 представлены кривые зависимости
коэффициента
![]() от коэффициента промысловой смертности
от коэффициента промысловой смертности![]() при размерах ячеи
при размерах ячеи![]() мм.
И здесь хорошо просматривается рост
коэффициента
мм.
И здесь хорошо просматривается рост
коэффициента![]() при увеличении
при увеличении![]() .Как и на рис. 7.6,
.Как и на рис. 7.6,![]() достигает наибольших значений при
достигает наибольших значений при![]() и
и![]() мм.
Такие же максимальные значения
мм.
Такие же максимальные значения![]() можно получить при совместном увеличении
коэффициента промысловой смертности
и размера ячеи.
можно получить при совместном увеличении
коэффициента промысловой смертности
и размера ячеи.

Рис. 7.6. Зависимость
коэффициента использования биомассы
поколения тихоокеанского хека от
![]() при коэффициентах промысловой
смертности
при коэффициентах промысловой
смертности![]() :
1–0,4; 2–0,6; 3–0,8; 4–1,0; 5–1,2; 6–1,4; 7–1,6.
:
1–0,4; 2–0,6; 3–0,8; 4–1,0; 5–1,2; 6–1,4; 7–1,6.
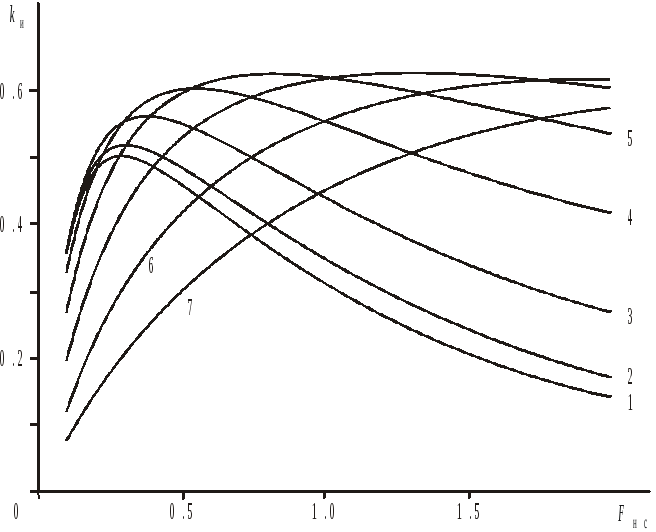
Рис. 7.7. Зависимость
коэффициента использования биомассы
поколения тихоокеанского хека от
![]() для размеров ячеи
для размеров ячеи![]() (мм): 1–50; 2–60; 3–70; 4–80; 5–90; 6–100; 7–110.
(мм): 1–50; 2–60; 3–70; 4–80; 5–90; 6–100; 7–110.
По
результатам сравнения полученных
зависимостей максимальное значение
коэффициента использования на примере
тихоокеанского хека получают при размере
ячеи в среднем 90мм. Коэффициент промысловой
смертности при условно неселективном
лове находится в пределах 0,7–1,4. Так как
для всех значений
![]() из этого интервала коэффициент
использования примерно одинаков, то
оптимальной интенсивности промысла
соответствует, очевидно,
из этого интервала коэффициент
использования примерно одинаков, то
оптимальной интенсивности промысла
соответствует, очевидно,![]() .
.
Абсолютный
максимум коэффициент использования
биомассы поколения, равный 0,62, имеет
при
![]() мм
и
мм
и![]() .
.
Для
улучшения промысловых и экономических
показателей лова иногда полезно уменьшить
![]() (промысловое усилие) и
(промысловое усилие) и![]() даже при снижении
даже при снижении![]() .
Так, в нашем примере допустимо принять
.
Так, в нашем примере допустимо принять![]() и
и![]() мм,
когда
мм,
когда![]() .
.
В
заключение отметим, что улов на единицу
пополнения
![]() и коэффициент
и коэффициент![]() отличаются лишь постоянным множителем.
Им является величина, обратная максимуму
биомассы
отличаются лишь постоянным множителем.
Им является величина, обратная максимуму
биомассы![]() .
Таким образом, обе характеристики
эффективности рыболовства пропорциональны.
.
Таким образом, обе характеристики
эффективности рыболовства пропорциональны.