

2.3. Статистические методы оценки пополнения промыслового стада
2.3.1. Рассмотренные в п. 2.2 предпосылки служат основой для разработки статистической оценки влияния различных факторов на пополнение промыслового страда.
2.3.2. Первый статистический метод основан на применении дисперсионного и корреляционного анализа для оценки степени влияния различных факторов на пополнение промыслового стада. Для этого используют данные многолетних наблюдений о зависимости пополнения от величины нерестового запаса.
Первоначально методами дисперсионного анализа оценивают степень связи нерестового запаса и пополнения по критерию Фишера при доверительной вероятности 0.90-0.95. Обычно связь устанавливают для нескольких градаций численности стада, например, малой, средней и высокой, т.к. влияние часто зависит от такой численности.
По результатам выполненного анализа, численность нерестового запаса обычно существенно влияет на пополнение. Это влияние заметно, если для отдельных интервалов численности нерестового запаса определить средние значения пополнения. Внутри каждого интервала численности нерестового запаса разброс значений пополнения, как правило, велик. Поэтому устанавливают силу связи между рассматриваемыми факторами отдельно для различных градаций численности нерестового запаса. Степень связи оценивают методами корреляционного анализа по критерию Стьюдента обычно с доверительной вероятностью 0,9-0,95.
Как правило, корреляционный анализ свидетельствуют, что для различных популяций и различных градаций численности нерестового запаса влияние посторонних факторов (в основном факторов внешней среды) на пополнение так же существенно, как и влияние численности нерестового запаса.
Обычно объем выборок для оценки влияния различных факторов на пополнение недостаточен для определения относительного влияния величины нерестового запаса и других факторов на пополнение. Но, очевидно, что влияние неодинаково для различных популяций и уровней численности нерестового запаса.
2.3.3. Второй статистический способ предусматривает разработку модели пополнения с учетом численности нерестового запаса, нерестовых площадей, количества корма и хищников, температуры воды.
Будем
считать, что при значениях всех факторов,
близких к оптимальным значениям,
наибольшая величина пополнения
![]() прямо пропорциональна численности
нерестового запаса
прямо пропорциональна численности
нерестового запаса![]() :
:
![]() , (2.3)
, (2.3)
где
![]() - эмпирический коэффициент.
- эмпирический коэффициент.
Приближенно
линию, соответствующую выражению (2.3),
проводят по экспериментальным данным
зависимости пополнения от нерестового
запаса через крайние левые точки (рис.
2.4). После построения прямой определяют
эмпирический коэффициент
![]() .
.
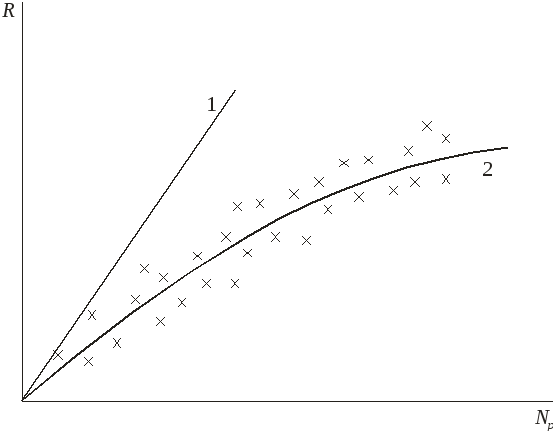
Рис. 2.4. Теоретическая (1) и реальная (2) кривая зависимости пополнения от величины нерестового запаса.
В реальных условиях при той же величине нерестового запаса пополнение меньше.
Обычно зависимость пополнения от влияющих факторов подчиняется экспоненциальным зависимостям или близка к закону нормального распределения. С учетом влияния всех факторов:
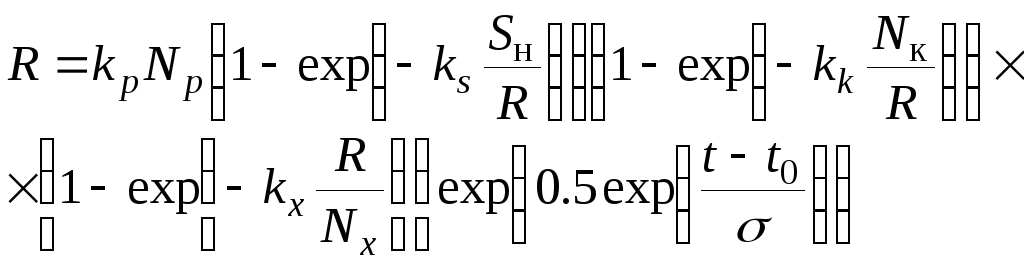 , (2.4)
, (2.4)
где
![]() ,
,![]() ,
,
![]() - эмпирические коэффициенты;
- эмпирические коэффициенты;![]() – нерестовые площади;
– нерестовые площади;![]() - количество корма;
- количество корма;![]() - количество хищников;
- количество хищников;![]() - температура воды;
- температура воды;![]() - оптимальная температура воды;
- оптимальная температура воды;![]() - среднеквадратичное отклонение по
температуре воды.
- среднеквадратичное отклонение по
температуре воды.
После оценки эмпирических коэффициентов
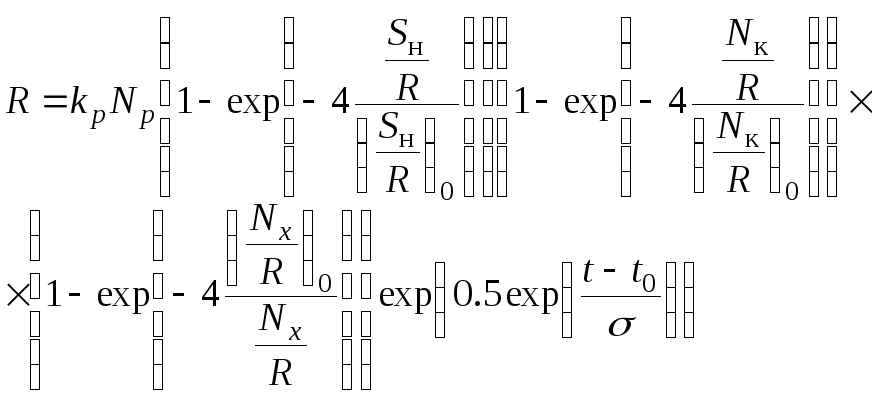 , (2.5)
, (2.5)
где
![]() - площадь нерестовых площадей на единицу
пополнения, при которой нерестовые
площади практически не влияют на величину
пополнения;
- площадь нерестовых площадей на единицу
пополнения, при которой нерестовые
площади практически не влияют на величину
пополнения;![]() - количество корма на единицу пополнения,
при котором количество корма достаточно
и не влияет на величину пополнения;
- количество корма на единицу пополнения,
при котором количество корма достаточно
и не влияет на величину пополнения;![]() - количество хищников на единицу
пополнения, при котором они практически
не влияют на численность пополнения.
- количество хищников на единицу
пополнения, при котором они практически
не влияют на численность пополнения.
В
правой части выражений (2.4) и (2.5) величина
![]() условно принята мерой плотности
концентрации рыб до вступления в
пополнение.
условно принята мерой плотности
концентрации рыб до вступления в
пополнение.
В соответствии с выражением (2.5) получим, в общем, нелинейную зависимость пополнения от численности нерестового стада для конкретных условий естественного воспроизводства.
При необходимости с помощью выражения (2.5) можно оценить степень влияния на пополнение каждого из факторов, считая значения других факторов оптимальными.
Формула (2.5) служит не столько для практического использования, сколько иллюстрацией второго способа оценки пополнения.
2.3.4. Третий способ оценки и прогнозирования пополнения промыслового стада основан на использовании метода группового учета аргументов (МГУА), предложенный первоначально для решения задач технической кибернетики (см. гл. 12)
Обычно при использовании МГУА пополнение определяют в функции величины промыслового стада, температуры воды во время нереста и в период роста молоди, состояния кормовой базы водоема, солености воды, количества хищников и других факторов, которые прямо или косвенно влияют на величину пополнения. При этом используют соответствующий статистический материал за 10-15 лет. Выполненные исследования, в частности, для каспийской кильки, показали преобладающее влияние температуры воды во время нереста на пополнение промыслового стада.
2.3.5. Кроме величины пополнения промыслового стада, необходимо знать возраст вступления пополнения в промысловое стадо и возраст вступления пополнения в промысел. Иногда предполагают одновременное вступление всех рыб одного поколения в промысловое стадо и одновременное вступление всех рыб этого поколения в промысел. Но обычно рыбы одного поколения вступают в промысловое стадо в течение нескольких лет, а при селективном лове рыбы различного размера и возраста не одновременно вступают в промысел.
Пополнение рыб одного поколения в промысловое стадо по возрасту и длине рыбы часто считают распределенным по закону нормального распределения. При этом начало кривой распределения часто считают соответствующим минимальному возрасту или росту рыб в облавливаемых скоплениях. Конец кривой приближенно увязывают с возрастом или размером рыб, соответствующим вершине кривой возрастного или размерного состава облавливаемых скоплений. Если известен состав улова, а не состав облавливаемых скоплений, то делают перерасчет с учетом кривой селективности орудия лова.
Кривые распределения пополнения и кривые селективности обычно частично накладываются друг на друга. При этом степень наложения во многом зависит от размера ячеи. При изменении размера ячеи кривая селективности смещается вправо или влево. Таким образом, при использовании функции кривой селективности в модели не вводят осредненный возраст вступления рыбы в промысловое стадо или возраст вступления рыбы в промысел. В необходимых случаях в качестве одного из пределов интегрирования используют минимальный возраст вступления рыбы в промысловое стадо.
Иногда все-таки необходимо использовать фиксированные значения двух рассматриваемых возрастов или длин рыб (вступления рыбы в промысловое стадо и в промысел). Тогда, например, за возраст вступления в промысловое стадо принимают его значение, соответствующее вершине кривой распределения пополнения, а за возраст вступления в промысловое стадо – значение ординаты кривой возрастной селективности равной 0.5.